
ABSTRAK
Famili gen patogenesis-related 1 (PR-1) sangat penting untuk pertahanan tanaman dan respons terhadap stres. Dalam penelitian ini, 11 gen SmPR-1 diidentifikasi dalam Salvia miltiorrhiza melalui analisis genomik yang komprehensif, yang semuanya mengkodekan protein dengan domain CAP (protein sekretori kaya sistein, antigen 5, dan patogenesis-related 1) yang terkonservasi dan peptida sinyal. Analisis filogenetik mengkategorikan gen-gen ini ke dalam lima kelompok evolusi, yang mencerminkan divergensi evolusinya. Analisis lokalisasi kromosom mengungkapkan bahwa famili gen SmPR-1 didistribusikan melintasi tiga kromosom: Chr1 berisi enam gen, Chr6 berisi tiga, dan Chr8 berisi satu. Analisis kolinearitas intraspesifik menunjukkan duplikasi segmental SmPR-1-5 dan SmPR-1-11 pada Chr1. Analisis kolinearitas interspesifik menunjukkan bahwa lima gen SmPR-1 kolinear dengan Arabidopsis thaliana dan Scutellaria baicalensis , dengan SmPR-1-1 juga menunjukkan kolinearitas dengan Oryza sativa dan Zea mays . Profil ekspresi spesifik jaringan menunjukkan tingkat ekspresi yang tinggi pada bunga dan batang, yang menunjukkan peran mereka dalam berbagai proses perkembangan. Pola ekspresi diferensial di bawah stres hormonal dan biotik mengungkapkan bahwa SmPR-1-5 sangat responsif terhadap pengobatan brassinosteroid (BR). Analisis lokalisasi subseluler menunjukkan bahwa SmPR-1-5 hadir dalam sitoplasma dan nukleus, yang menunjukkan keterlibatannya dalam pensinyalan intraseluler. Selain itu, infeksi CMV memicu pola ekspresi yang bergantung waktu, mengaktifkan gen spesifik selama tahap infeksi awal dan akhir. Temuan ini memberikan wawasan berharga tentang peran fungsional gen SmPR-1 dalam respons stres dan imunitas, yang meletakkan dasar untuk pemuliaan varietas S. miltiorrhiza yang tahan penyakit . Penelitian di masa mendatang harus mengeksplorasi mekanisme pengaturan dan interaksi gen SmPR-1 dengan jalur pertahanan lain untuk sepenuhnya memahami kontribusinya terhadap ketahanan tanaman.
1 Pendahuluan
Respon imun tanaman adalah sistem multifaset yang dirancang untuk mengurangi efek berbahaya dari mikroorganisme patogen dan mencegah infeksi dan kerusakan jaringan. Ini terdiri dari dua strategi pertahanan utama: kekebalan berbasis pengenalan pola (PRBI), yang merupakan garis pertahanan pertama, dan kekebalan yang dipicu oleh efektor (ETI), yang menyajikan respons yang lebih spesifik dan kuat. Selain itu, gen terkait patogenesis (PR) yang mengkode protein PR bertindak secara sinergis untuk meningkatkan ketahanan tanaman terhadap invasi patogen. Interaksi sinergis antara mekanisme ini memungkinkan tanaman untuk memasang respons imun yang tahan lama dan kuat (Silva et al. 2018 ; Takken dan Tameling 2009 ; Wan et al. 2019 ). Gen terkait patogenesis (PR) mewakili kelompok elemen genetik yang berbeda yang diaktifkan pada tanaman selama pertemuan patogen dan paparan stresor abiotik. Gen-gen ini memainkan peran penting dalam pertahanan tanaman dengan memulai kaskade perlindungan dan mengkode protein PR yang penting untuk ketahanan terhadap penyakit (Hamamouch et al. 2010 ; Punja 2001 ; Sung et al. 2021 ). Gen-gen ini tidak hanya diekspresikan selama infeksi mikroba tetapi juga berkontribusi pada proses-proses penting tanaman, termasuk pematangan, pembungaan, plasmolisis, dan penuaan (Breen et al. 2017 ; Zribi et al. 2021 ). Protein PR meningkatkan toleransi tanaman terhadap patogen dengan mengakumulasi berbagai peptida dan protein (Punja 2001 ). Selain itu, mereka dapat merespons berbagai stresor abiotik, termasuk kekeringan, pembekuan, radiasi ultraviolet, salinitas, stres osmotik, dan sinyal hormonal seperti asam salisilat (SA), asam absisat (ABA), asam jasmonat (JA), dan auksin (IAA) (Agrawal et al. 2000 ; Akbudak et al. 2020 ; AlHudaib et al. 2022 ; Ghorbel et al. 2021 ; Mou et al. 2003 ). Dicirikan oleh ukuran molekulnya yang kecil, stabilitas panas, dan ketahanan terhadap protease, protein PR diekspresikan secara luas dalam berbagai jaringan tanaman (van Loon et al. 1994 ). Pertama kali ditemukan pada tahun 1970-an pada tanaman tembakau yang terinfeksi virus mosaik tembakau (TMV) dan diinduksi oleh asam salisilat atau asetilsalisilat (Van Loon dan Van Kammen 1970 ), keluarga protein PR sejak itu telah berkembang menjadi 17 kelompok berbeda berdasarkan kesamaan dalam urutan asam amino, hubungan imunologis, dan fungsi katalitik (Sels et al. 2008 ).
Keluarga protein PR-1 adalah protein terkait patogenesis yang pertama kali diidentifikasi dan tersebar luas pada tumbuhan monokotil dan dikotil (Akbudak et al. 2020 ; Fraser 1981 ; Kothari et al. 2016 ; Lawrence et al. 1996 ; Li et al. 2023 ; Shin et al. 2014 ; Zhang et al. 2022a ). Terkenal karena peran pentingnya dalam resistensi yang diperoleh secara sistemik (SAR), PR-1 menandai status pertahanan tanaman dengan memfasilitasi kematian sel terprogram (Breen et al. 2017 ; Chassot et al. 2007 ). Sangat terkonservasi di seluruh spesies tanaman, protein PR-1 diklasifikasikan menjadi tipe asam dan basa berdasarkan titik isoelektriknya (Breen et al. 2017 ). Protein-protein ini melakukan fungsi ganda: mereka disekresikan ke dalam ruang ekstraseluler atau disimpan dalam vakuola (Sessa et al. 1995 ). Fitur penentu protein PR-1 adalah domain CAP (cysteine-rich secretory protein, antigen 5, and pathogenesis-related 1) yang terkonservasi, yang terdiri dari sekitar 150 residu asam amino yang membentuk struktur stabil dari empat α-heliks dan empat β-untai yang diperkuat oleh ikatan disulfida (Breen et al. 2017 ). Domain CAP memberikan aktivitas antimikroba dengan mengenali dan mengikat sterol pada membran patogen dan menghambat pertumbuhannya (Chen et al. 2014 ; Schneiter and Di Pietro 2013 ). Elemen struktural ini penting untuk peran fisiologis PR-1 dalam melawan stres biologis dan lingkungan (Ghorbel et al. 2021 ). Akumulasi protein PR-1 dalam jaringan tanaman sangat terkait dengan infeksi patogen. Misalnya, pada daun tembakau yang terinfeksi TMV, protein PR-1 mencakup 1%–2% dari total protein daun (Alexander et al. 1993 ). Pada tomat, PR-1c menekan perkecambahan spora dan pertumbuhan tabung kecambah Phytophthora infestans , mengurangi kejadian penyakit (Niderman et al. 1995 ). Temuan ini menunjukkan bahwa protein PR-1 adalah perantara penting dalam pertahanan tanaman. Pada tanaman teh, gen PR-1 merespons secara aktif terhadap stres akibat penyakit hawar blister, yang terkait erat dengan jalur pensinyalan utama, termasuk siklus TCA, pensinyalan yang dimediasi NPR1, EDS16, BGL2, PR4, dan HCHIB (Zhang et al. 2022a ).
Gen PR-1 sangat penting untuk kekebalan tanaman terhadap penyakit dan memainkan peran penting dalam manajemen stres abiotik. Pada gandum, gen TaPR-1-1 dapat diinduksi oleh stres dingin, garam, dan osmotik, dengan ekspresinya meningkatkan toleransi stres pada ragi dan Arabidopsis (Wang et al. 2019 ). Pada tomat, PR-1b1 meningkat pada suhu rendah, dan ekspresinya yang berlebihan pada tanaman transgenik meningkatkan adaptasi stres (Goyal et al. 2016 ; Kiba et al. 2007 ; Sarowar et al. 2005 ). Pada padi, protein OsSAP1 mengaktifkan gen terkait stres endogen, termasuk OsAMTR1, SCP/TAPS, dan protein terkait patogenesis OsSCP (Kothari et al. 2016 ). Dalam Arabidopsis , pengatur transkripsi yang diinduksi kekeringan Di19 meningkatkan ekspresi gen-gen yang terkait dengan patogenesis, yang selanjutnya memperkuat ketahanan terhadap stres (Liu et al. 2013 ). Faktor transkripsi NTL6, anggota keluarga NAC, secara langsung mengikat ke daerah promotor gen-gen yang terkait dengan patogenesis yang responsif terhadap dingin, seperti PR-1, PR-2, dan PR-5, yang merangsang ekspresinya (Seo et al. 2010 ). Gen-gen PR ini diatur oleh stres abiotik dan juga oleh hormon tanaman dan penginduksi kimia, termasuk ET, SA, JA, brassinosteroid (BR), benzothiadiazole (BTH), dan asam isonicotinic (INA) (Breen et al. 2017 ; Chandrashekar et al. 2018 ; Edreva dan Kostoff 2005 ; Van Loon dan Van Strien 1999 ; Zhang et al. 2015 ). Singkatnya, gen PR-1 memainkan peran penting dalam respons tanaman terhadap berbagai tekanan biotik dan abiotik, dan transkripsinya dikontrol ketat oleh berbagai faktor pengatur.
Salvia miltiorrhiza adalah herba menahun dari genus Salvia dalam famili Lamiaceae yang berfungsi sebagai komponen berharga dari pengobatan tradisional Tiongkok, dan akarnya umumnya digunakan untuk tujuan pengobatan (Sui 2019 ). Terkenal untuk mengobati penyakit kardiovaskular, ia meningkatkan sirkulasi darah, mengatur menstruasi, menenangkan pikiran, dan mengurangi pembengkakan dan nyeri (Wu dan Wang 2012 ). Umurnya yang pendek, pertumbuhan yang kuat, teknik modifikasi genetika yang canggih, genom yang kompak, dan jumlah kromosom yang kecil menjadikan S. miltiorrhiza sebagai model yang ideal untuk penelitian tanaman farmasi (Boli 2009 ; Xu et al. 2016 ; Zhang et al. 2015 ). Dibudidayakan secara luas di Tiongkok, area penanamannya yang terus bertambah semakin terancam oleh penyakit, termasuk infeksi jamur, yang memengaruhi hasil dan kualitas (Lu et al. 2021 ; Wang et al. 2018 ; Zhang et al. 2022b ). Meskipun peran penting keluarga gen PR-1 dalam ketahanan tanaman terhadap penyakit, fungsi spesifiknya dalam S. miltiorrhiza masih belum cukup dieksplorasi. Dalam penelitian ini, kami mengkarakterisasi keluarga gen SmPR-1 menggunakan data genomik dari S. miltiorrhiza dan melakukan analisis bioinformatika yang mendalam. Melalui analisis transkriptom dan PCR kuantitatif waktu nyata (qRT-PCR), kami memeriksa profil ekspresi gen SmPR-1 dalam infeksi virus mosaik mentimun dan perlakuan hormon tanaman untuk mengidentifikasi gen dengan peran fungsional yang signifikan. Penelitian ini memberikan landasan teoritis untuk penelitian lebih lanjut tentang fungsi biologis keluarga gen SmPR-1 dan menyarankan strategi baru untuk manajemen penyakit dan peningkatan kualitas tanaman obat ini.
2 Hasil
2.1 Identifikasi Anggota Keluarga Gen SmPR-1
Dalam genom S. miltiorrhiza , 11 sekuens SmPR-1 diidentifikasi menggunakan alat NCBI Batch CD-Search dan ditetapkan sebagai SmPR-1-1 hingga SmPR-1-11 . Sekuens ini mengandung domain CAP, yang merupakan karakteristik protein sekretori kaya sistein, antigen komponen 5, dan protein PR-1. Panjang sekuens bervariasi, dengan SmPR-1-8 menjadi yang terpanjang dengan 210 asam amino, sedangkan SmPR-1-5, SmPR-1-9, dan SmPR-1-10 adalah yang terpendek dengan 159 asam amino. Mayoritas (90,91%) sekuens berkisar antara 150 hingga 200 asam amino. Berat molekul (BM) protein ini berkisar antara 17,44 hingga 23,74 kDa, dan titik isoelektrik teoritis (PI) mereka bervariasi dari 4,56 hingga 9,8 (Tabel 1 ). Semua protein SmPR-1 diidentifikasi sebagai hidrofilik, sebagaimana ditunjukkan oleh indeks GRAVY negatifnya, dan peptida sinyal dideteksi untuk setiap urutan. Penyelarasan urutan menunjukkan kesamaan yang tinggi antara protein SmPR-1. Prediksi lokalisasi subseluler menunjukkan bahwa protein SmPR-1 sebagian besar terletak di kloroplas dan ruang ekstraseluler, yang konsisten dengan pengamatan pada spesies tanaman lain (Tabel S1 ).
| ID Urutan | Gen | Panjang protein (aa) | Berat jenis (kDa) | saya | SAUS | Pencarian HMM nilai- E | Lokasi subseluler | Bahasa Inggris | Panjang CDS | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SMILT018280.1 | SmPR-1-1 | 165 | Jam 18.30 | 9.64 | -0,278 | 6.30E-22 | Kloroplas | 26–27 | 498 | ||
| SMILT018281.1 | SmPR-1-2 | 188 | 20.46 | 6.72 | -0,08 | 2.00E-19 | Kloroplas | 24–25 | 567 | ||
| SMILT008035.1 | SmPR-1-3 | 167 | 18.62 | 8.97 | -0,18 | 1.20E-20 | Kloroplas | 25–26 | 504 | ||
| SMILT009945.1 | SmPR-1-4 | 170 | 18.52 | 8.38 | -0,252 | 5.10E-16 | Kloroplas | 22–23 | 513 | ||
| SMILT013753.1 | SmPR-1-5 | 159 | 17.68 | 8.27 | -0,29 | 6.60E-20 | Ekstrasel | 22–23 | 480 | ||
| SMILT013754.1 | SmPR-1-6 | 161 | tanggal 18.12 | Tanggal 9.04 | -0,466 | 2.30E-20 | Ekstrasel | 24–25 | 486 | ||
| SMILT013755.1 | SmPR-1-7 | 161 | 17.44 | 6.64 | -0,207 | 7.70E-24 | Ekstrasel | 23–24 | 486 | ||
| SMILT014473.1 | SmPR-1-8 | 210 | 23.74 | 9.8 | -0,358 | 1.60E-26 | Kloroplas | 26–27 | 633 | ||
| SMILT015244.1 | SmPR-1-9 | 159 | 17.76 | 5.51 | -0,377 | 1.30E-21 | Ekstrasel | 21–22 | 480 | ||
| SMILT015245.1 | SmPR-1-10 | 159 | 17.76 | 4.56 | -0,393 | 1.70E-20 | Kloroplas | 24–25 | 480 | ||
| SMILT015247.1 | SmPR-1-11 | 164 | 18.38 | 5.16 | -0,336 | 9.00E-23 | Ekstrasel | tahun 20–21 | 495 | ||
2.2 Perbandingan Urutan Alignment dan Analisis Filogenetik Protein SmPR-1
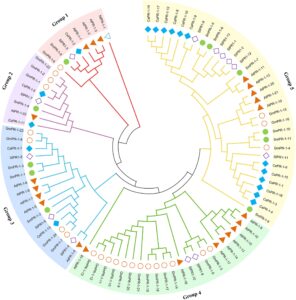
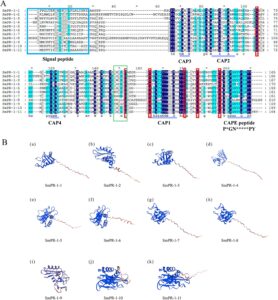
Analisis filogenetik gen PR-1 dari Salvia miltiorrhiza (Sm), Arabidopsis thaliana (At), Camellia sinensis (Cs), Solanum lycopersicum (Sl), Glycine max (Gm), dan Nicotiana tabacum (Nt) mengungkapkan hubungan evolusi yang signifikan dan divergensi fungsional dalam keluarga gen ini. Di antara 88 sekuens PR-1 yang dianalisis, muncul lima kelompok berbeda, yang mewakili garis keturunan evolusi yang unik. Kelompok 1 dan 2 mengandung satu gen SmPR-1 ( SmPR-1 , SmPR-8 , dan SmPR-4 ), yang menyoroti status evolusi mereka yang relatif terisolasi. Kelompok 3, yang meliputi tiga gen SmPR-1 ( SmPR-1-1 , SmPR-2 , dan SmPR-3 ), menunjukkan potensi redundansi fungsional atau spesialisasi di antara anggota yang terkait erat. Kelompok 5 adalah kelompok terbesar, termasuk enam gen SmPR-1 ( SmPR-1-5 , SmPR-6 , SmPR-7 , SmPR-9 , SmPR-10 , dan SmPR-11 ), mungkin menunjukkan peristiwa duplikasi baru-baru ini dan diversifikasi berikutnya (Gambar 1 ). Penyelarasan sekuens mengungkapkan bahwa sebagian besar protein SmPR-1 memiliki enam residu sistein yang terkonservasi yang penting untuk stabilitas struktural dan resistensi patogen. Namun, peptida turunan CAP yang tidak lengkap (CAPE) dalam SmPR-1-4 dan SmPR-1-8 menunjukkan bahwa modifikasi evolusioner yang unik berpotensi memengaruhi interaksi patogenik. Selain itu, Kelompok 3 dan 5 membentuk kluster filogenetik berbeda yang dicirikan oleh kesamaan intragroup yang tinggi dan lintasan evolusi yang berbeda (Gambar 2A ), yang berpotensi mencerminkan spesialisasi fungsional atau adaptasi terhadap tantangan lingkungan tertentu. Prediksi struktur tiga dimensi dari 11 protein SmPR-1 mengungkapkan bahwa protein dari gen SmPR-1 yang secara filogenetik dekat cenderung memiliki struktur yang lebih mirip, terutama yang berada dalam kelompok yang sama. Misalnya, SmPR-1-10 dan SmPR-1-11 dalam Kelompok 5 menunjukkan kesamaan struktural yang tinggi (Gambar 2B ), yang konsisten dengan hasil dari analisis motif dan analisis struktur gen.
2.3 Analisis Lokalisasi dan Kolinearitas Kromosom SmPR-1
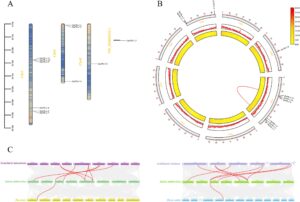
Analisis lokalisasi kromosom mengungkapkan bahwa di antara delapan kromosom, hanya tiga yang mengandung gen SmPR-1 : enam pada Chr1, tiga pada Chr6, dan satu pada Chr8, dengan satu sekuens yang tidak dirakit (Gambar 3A ). Analisis kolinearitas intraspesifik menunjukkan bahwa di antara 11 gen SmPR-1 , hanya SmPR-1-5 dan SmPR-1-11 yang menunjukkan duplikasi segmental, yang keduanya terletak pada Chr1 (Gambar 3B ). Analisis rasio mutasi sinonim dan nonsinonim (Ka/Ks) dari segmen yang terduplikasi mengungkapkan nilai Ka/Ks kurang dari 1, yang menunjukkan bahwa gen-gen ini mungkin telah mengalami seleksi pemurnian. Analisis kolinearitas interspesifik dari keluarga gen SmPR-1 dengan A. thaliana , Scutellaria baicalensis , Oryza sativa , dan Zea mays mengungkapkan bahwa keluarga gen SmPR-1 kolinear dengan keempat spesies. Analisis lebih lanjut menunjukkan bahwa jumlah tertinggi pasangan gen kolinear terjadi pada A. thaliana dan S. baicalensis (masing-masing delapan pasang), sedangkan yang terendah diamati pada O. sativa dan Z. mays (masing-masing satu pasang). Khususnya, lima gen SmPR-1 (S mPR-1-1 , SmPR-1-4 , SmPR-1-5 , SmPR-1-8 , dan SmPR-1-11 ) menunjukkan kolinearitas interspesifik dengan A. thaliana dan S. baicalensis , dan satu gen SmPR-1 ( SmPR-1-1 ) menunjukkan kolinearitas interspesifik dengan keempat spesies (Gambar 3C ).
2.4 Analisis Motif Struktural dan Konservasi SmPR-1
Analisis struktural dan motif gen SmPR-1 dilakukan menggunakan perangkat lunak TBtools untuk mendapatkan wawasan lebih dalam tentang karakteristiknya. Hasil prediksi motif mengidentifikasi motif 1, 2, 3, dan 4 di semua gen, yang tersusun secara konsisten, menunjukkan konservasi tinggi dan kemungkinan perannya sebagai domain CAP. Motif 5 dan 6 juga relatif terkonservasi dan muncul di sebagian besar sekuens. Motif unik diidentifikasi: motif 7 eksklusif untuk SmPR-1-4 dan SmPR-1-8, motif 10 untuk SmPR-1-2 dan SmPR-1-4, dan motif 9 untuk SmPR-1-4 dan SmPR-1-8. Analisis sekuens DNA mengungkapkan bahwa semua gen SmPR-1 mengandung satu ekson dengan panjang yang sama, tanpa intron yang ada pada gen mana pun (Gambar 4 ).
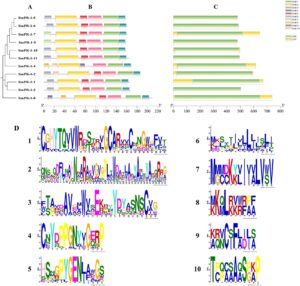
Bahasa Indonesia: Untuk lebih memahami keluarga gen SmPR-1 di S. miltiorrhiza , kami melakukan analisis terperinci dari struktur gen dan komposisi motif menggunakan perangkat lunak TBtools. Analisis mengidentifikasi 10 motif berbeda (motif 1–10) dalam keluarga gen SmPR-1 . Khususnya, motif 1, 2, 3, dan 4 secara konsisten hadir di semua gen SmPR-1 dalam susunan yang seragam, yang menunjukkan tingkat konservasi yang tinggi dan menunjukkan bahwa peran mereka sebagai komponen inti dari domain CAP yang dilestarikan sangat penting untuk fungsi protein PR-1 (Gambar 3 ). Motif 5 dan 6 diidentifikasi sebagai relatif dilestarikan dan hadir di sebagian besar urutan SmPR-1 , mendukung hipotesis konservasi fungsional dalam keluarga gen ini (Gambar 3C ). Sebaliknya, motif tertentu menunjukkan distribusi terbatas, yang menunjukkan potensi divergensi fungsional. Misalnya, motif 7 unik untuk SmPR-1-4 dan SmPR-1-8 ; motif 10 hanya muncul di SmPR-1-2 dan SmPR-1-4 ; dan motif 9 eksklusif untuk SmPR-1-4 dan SmPR-1-8 (Gambar 4B,D ). Pola motif unik ini mungkin menunjukkan peran khusus dalam menanggapi isyarat lingkungan tertentu atau proses perkembangan. Analisis struktural mengungkapkan bahwa semua gen SmPR-1 memiliki satu ekson tanpa intron, dan panjang ekson relatif seragam di seluruh famili (Gambar 4C ). Struktur ekson tunggal yang dilestarikan ini selaras dengan gen PR-1 lainnya pada spesies tanaman lain, yang menunjukkan konservasi evolusioner dan efisiensi regulasi potensial.
2.5 Prediksi Elemen Cis -Acting dan Faktor Transkripsi di Antara Gen SmPR-1
Analisis elemen pengatur cis (CRE) dalam urutan promotor gen SmPR-1 mengidentifikasi beragam CRE, yang dikategorikan ke dalam lima kelas fungsional: elemen terkait transkripsi umum (Grup 1), elemen responsif hormon (Grup 2), elemen responsif stres (Grup 3), elemen terkait pertumbuhan dan perkembangan (Grup 4), dan elemen responsif cahaya (Grup 5). Beberapa elemen responsif hormon dideteksi, termasuk yang responsif terhadap SA (TCA, as-1, dan elemen TCA), IAA (elemen AuxRE dan TGA), ET (ERE), giberelin (GA) (motif CARE, GARE, p-box, dan TATC box), metil jasmonat (MeJA) (motif TGACG dan motif CGTCA), dan ABA (ABRE, ABRE4, ABRE3a, motif AAAGAA, dan inti DRE) (Gambar 5 ). CRE terkait SA dan MeJA, yang sangat terkait dengan mekanisme pertahanan, sangat melimpah di sebagian besar gen SmPR-1 . Selain itu, elemen responsif stres, seperti situs pengikatan MYB dan kotak W, menonjol, yang menunjukkan potensi keterlibatan gen SmPR-1 dalam respons stres biotik dan abiotik.

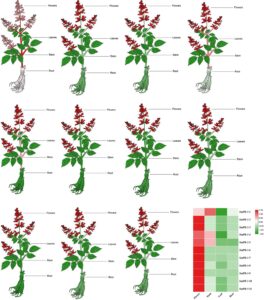
2.6 Profil Transkripsi Gen SmPR-1 di Berbagai Jaringan
Pola ekspresi gen SmPR-1 di S. miltiorrhiza mengungkapkan karakteristik spesifik organ yang berbeda, yang menunjukkan potensi divergensi fungsional. Di antara 11 gen SmPR-1 , SmPR-1-1 , SmPR-1-4 , dan SmPR-1-9 menunjukkan tingkat ekspresi yang sangat tinggi pada bunga, yang menunjukkan kemungkinan peran mereka dalam perkembangan bunga dan proses reproduksi. Selain itu, SmPR-1-1 dan SmPR-1-9 diekspresikan secara signifikan di batang, yang menunjukkan keterlibatan mereka dalam proses biologis spesifik dalam jaringan ini. Sebagian besar gen SmPR-1 menunjukkan tingkat ekspresi rendah pada daun dan akar, kecuali SmPR-1-1 , yang menunjukkan ekspresi sedang pada akar (Gambar 6 ). Temuan ini menunjukkan fungsi yang beragam dari gen SmPR-1 , yang menekankan ekspresi spesifik organ mereka pada bunga, batang, dan akar.
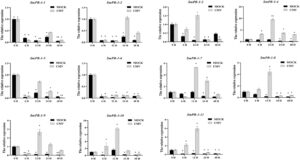
2.7 Respon Transkripsi Gen SmPR-1 terhadap Infeksi CMV
Analisis qRT-PCR mengevaluasi profil transkripsi gen SmPR-1 di S. miltiorrhiza setelah infeksi virus mosaik mentimun (CMV) pada lima titik waktu: 0, 6, 12, 24, dan 48 jam pascainfeksi. Hasil ini mengungkapkan pola ekspresi yang berbeda di seluruh keluarga gen SmPR-1 . Khususnya, SmPR-1-4 , SmPR-1-7 , dan SmPR-1-9 menunjukkan peningkatan ekspresi yang signifikan pada 12 dan 24 jam pascainfeksi, dengan peningkatan kadar yang bertahan pada 48 jam (Gambar 7 ), yang menunjukkan keterlibatan mereka dalam respons pertahanan berkelanjutan terhadap CMV. Sebaliknya, SmPR-1-10 dan SmPR-1-11 menunjukkan respons ekspresi awal dan cepat, memuncak pada 6 dan 12 jam, diikuti oleh penurunan pada 24 jam, yang menunjukkan peran mereka dalam pertahanan awal terhadap infeksi virus. Variasi ini menyoroti mekanisme pertahanan multifase yang kompleks terhadap CMV di S. miltiorrhiza .
2.8 Ekspresi Gen SmPR-1 yang Diinduksi Stres Hormonal
Identifikasi motif cis -regulatory di daerah promoter SmPR-1 telah mengungkap banyak elemen yang responsif terhadap hormon, yang menyoroti potensi keterlibatan gen SmPR-1 dalam jalur pensinyalan yang dimediasi hormon. Analisis transkriptom S. miltiorrhiza selanjutnya mengonfirmasi responsivitas gen SmPR-1 spesifik terhadap pengobatan asam absisat (ABA) dan SA, yang menunjukkan peran mereka dalam regulasi hormonal yang kompleks. Keluarga gen SmPR-1 menunjukkan respons yang bergantung pada waktu yang berbeda terhadap hormon-hormon ini. Di bawah pengaruh pengobatan ABA, gen SmPR-1-3 menunjukkan tren peningkatan regulasi yang signifikan setelah 8 jam pengobatan, sedangkan gen SmPR-1-5 menunjukkan peningkatan ekspresi setelah 2 jam pengobatan, mencapai puncaknya pada 4 jam. Sebaliknya, tingkat ekspresi gen SmPR-1-1 , SmPR-1-4 , dan SmPR-1-7 secara bertahap menurun setelah pengobatan, yang menunjukkan kemungkinan keterlibatan mereka dalam jalur pensinyalan ABA dan proses biologis terkait. Di bawah perlakuan SA, tingkat ekspresi gen SmPR-1-1 , SmPR-1-4 , dan SmPR-1-7 awalnya meningkat setelah 2 jam perlakuan SA tetapi kemudian menunjukkan tren menurun setelah 8 jam. Sebaliknya, tingkat ekspresi gen SmPR-1-2 , SmPR-1-3 , SmPR-1-5 , dan SmPR-1-10 terus menurun (Gambar 8A ). Temuan ini mengungkap perubahan ekspresi kompleks dan dinamis dari keluarga gen SmPR-1 di bawah induksi SA, yang menyoroti peran potensial mereka dalam respons pertahanan tanaman.
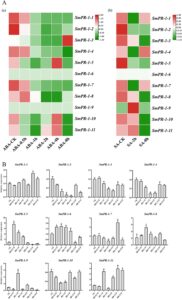
Respons transkripsi gen SmPR-1 terhadap pengobatan hormon BR mengungkapkan variasi temporal dan spesifik gen yang signifikan. SmPR-1-5 , SmPR-1-8 , dan SmPR-1-9 menunjukkan induksi cepat pada titik waktu awal (0,5 dan 1 jam), dengan SmPR-1-5 menampilkan tingkat ekspresi tertinggi pada 0,5 jam, diikuti oleh penurunan bertahap. Sebaliknya, SmPR-1-1 dan SmPR-1-7 menunjukkan peningkatan sedang dalam ekspresi pada titik waktu jangka menengah (6 jam), yang menurun pada tahap selanjutnya. Khususnya, SmPR-1-2 dan SmPR-1-4 menunjukkan ekspresi yang berkurang pada titik waktu awal tetapi secara bertahap meningkatkan ekspresi pada tahap selanjutnya (Gambar 8B ). Hasil-hasil ini menunjukkan respons spesifik dan diferensial gen SmPR-1 terhadap pengobatan BR, yang menunjukkan peran fungsional yang berbeda dalam proses biologis yang dimediasi BR.
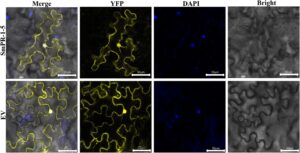
2.9 Analisis Lokalisasi Subseluler Protein SmPR-1
Analisis bioinformatika dari sekuens asam amino SmPR-1 memperkirakan bahwa sebagian besar protein SmPR-1 terlokalisasi di kloroplas atau ruang ekstraseluler. Di antara keluarga gen SmPR-1, SmPR-1-5 menunjukkan ekspresi terkuat dalam respons terhadap stres BR, seperti yang ditentukan oleh qRT-PCR, menjadikannya kandidat untuk studi lokalisasi subseluler terperinci menggunakan mikroskopi konfokal laser. DAPI digunakan sebagai pewarna nuklir, dan pHB-YFP berfungsi sebagai penanda fluoresensi. Hebatnya, hasil tersebut mengungkapkan bahwa SmPR-1-5 terlokalisasi tidak hanya pada membran sel tetapi juga di dalam nukleus (Gambar 9 ), melebihi ekspektasi awal. Lokalisasi ganda ini menunjukkan peran fungsional kompleks SmPR-1-5, yang mengintegrasikan pertahanan ekstraseluler dengan fungsi regulasi intraseluler. Temuan ini memberikan wawasan baru tentang fleksibilitas protein keluarga PR-1 dan menyoroti SmPR-1-5 sebagai mediator utama dari sinyal pertahanan dan regulasi gen di S. miltiorrhiza .
3 Diskusi
Protein PR-1 merupakan salah satu protein yang paling banyak disintesis selama respons imun tanaman, berfungsi sebagai indikator utama peningkatan sinyal pertahanan, dan memainkan peran penting dalam resistensi sistemik yang diperoleh (SAR) (van Loon et al. 2006 ; Van Loon dan Van Strien 1999 ). Kemajuan pesat dalam metodologi pengurutan genom yang komprehensif telah mendorong penyelidikan keluarga gen PR-1 di berbagai spesies tanaman (Akbudak et al. 2020 ; AlHudaib et al. 2022 ; Chu et al. 2022 ; van Loon et al. 2006 ; Lu et al. 2011 ; Yin et al. 2023 ; Zhang et al. 2022a ; Zribi et al. 2023 ). Penelitian telah menyoroti peran penting keluarga gen PR-1 dalam mekanisme pertahanan tanaman terhadap penyakit. Bahasa Indonesia: Salvia miltiorrhiza adalah herba menahun yang banyak digunakan dalam pengobatan tradisional dan rentan terhadap banyak hama dan penyakit. Ketergantungan pada pestisida kimia untuk pengendalian hama berisiko mendorong resistensi hama dan meninggalkan residu yang menurunkan kualitas bahan obat, sehingga memengaruhi kemanjuran terapeutik (Wang et al. 2018 ). Akibatnya, mengidentifikasi gen resistensi dalam genom S. miltiorrhiza dan mengembangkan varietas yang tahan penyakit sangat penting untuk meningkatkan nilai pengobatan dan kemanjuran klinisnya. Meskipun penting, studi molekuler pada gen PR-1 di S. miltiorrhiza masih belum dieksplorasi. Studi ini merupakan analisis genomik komprehensif pertama dari gen PR-1 di S. miltiorrhiza dengan tujuan untuk menjelaskan peran mereka dalam respons stres biotik dan abiotik.
Studi ini mengidentifikasi 11 protein PR-1 dalam genom S. miltiorrhiza , jumlah yang relatif rendah dibandingkan dengan yang ada di tumbuhan dikotil dan monokotil lainnya. Perbedaan ini mungkin terjadi karena redundansi fungsional di antara anggota PR-1, yang menunjukkan bahwa ukuran genom tidak berkorelasi langsung dengan jumlah gen PR-1 (AlHudaib et al. 2022 ). Analisis filogenetik dari famili gen PR-1 di tujuh spesies ( Arabidopsis thaliana , Camellia sinensis , Solanum lycopersicum , Glycine max , Nicotiana tabacum , Triticum aestivum , dan S. miltiorrhiza ) mengklasifikasikan 111 gen PR-1 ke dalam lima klade. 11 gen SmPR-1 didistribusikan di empat klade dengan motif yang dilestarikan, yang mencerminkan adaptasi evolusioner terhadap tekanan lingkungan tertentu. Khususnya, S. miltiorrhiza memiliki jumlah gen PR-1 tertinggi dalam klade 5 (Gambar 2 ), yang menunjukkan peran penting bagi kelompok ini dalam mekanisme pertahanan. Klasifikasi gen PR-1 dalam S. miltiorrhiza menunjukkan fitur yang sama dan unik dibandingkan dengan spesies lain. Misalnya, 23 gen OsPR-1 dalam beras telah dikelompokkan menjadi protein asam (Grup II) dan basa (Grup III) berdasarkan titik isoelektriknya (pI), dengan Grup II mengandung protein asam dan Grup III mengandung protein basa (Liu dan Xue 2006 ). Demikian pula, 23 gen TaPR-1 dalam gandum diklasifikasikan menjadi protein asam, basa, dan basa dengan ekstensi C-terminal (Lu et al. 2011 ), sementara gen PR-1 teh dikategorikan menjadi lima klade berdasarkan peptida sinyal, pI, dan ekstensi C-terminal (Zhang et al. 2022a ). Sebaliknya, gen PR-1 S. miltiorrhiza yang mengkode protein asam dan basa mengelompok dalam kelompok yang sama, serupa dengan temuan Akbudak et al. ( 2020 ) dan tebu (Chu et al. 2022 ), di mana protein PR-1 mengelompok terlepas dari sifat asam atau basanya.
Protein PR-1 secara umum diklasifikasikan ke dalam dua kelompok berdasarkan titik isoelektrik (pIs) mereka: asam dan basa (Kiraga et al. 2007 ). Di antara 11 protein PR-1 yang diidentifikasi dalam S. miltiorrhiza , sembilan bersifat basa dan dua bersifat asam. Protein PR-1 basa biasanya dikaitkan dengan aktivitas antimikroba yang kuat. Misalnya, dalam tomat, protein PR-1c dan PR-1g basa dapat menunjukkan aktivitas antimikotik yang lebih besar daripada protein PR-1a dan PR-1b yang bersifat asam (Niderman et al. 1995 ). Demikian pula, ekspresi berlebih gen PR-1 basa dari lada dalam tembakau secara signifikan meningkatkan resistensi terhadap stres logam berat dan bakteri patogen (Sarowar et al. 2005 ). Dalam teh, 10 dari 17 protein CsPR-1 bersifat basa dan diekspresikan dengan cepat selama infeksi penyakit busuk daun blister. Meskipun gen SmPR-1-10 dan SmPR-1-11 yang bersifat asam menunjukkan respons awal terhadap infeksi, ekspresinya menurun secara signifikan selama tahap selanjutnya (Gambar 6 ). Pola ekspresi yang bervariasi ini menunjukkan mekanisme pertahanan yang kompleks terhadap CMV. Khususnya, semua 11 gen SmPR-1 yang diidentifikasi dalam penelitian ini tidak memiliki intron (Gambar 1 dan 3 ), yang dapat memfasilitasi regulasi transkripsi cepat di bawah tekanan. Tidak adanya intron dihipotesiskan sebagai adaptasi evolusi yang memungkinkan respons yang lebih cepat terhadap tekanan lingkungan, seperti yang diamati dalam teh, di mana gen CsPR-1 yang tidak memiliki intron diekspresikan lebih cepat selama stres layu Fusarium (Zhang et al. 2022a ). Selain itu, SmPR-1-6 , SmPR-1-7 , SmPR-1-8 , dan SmPR-1-9 tidak memiliki motif pengikat caveolin (CBM), yang berpotensi mengganggu kemampuan mereka untuk mengikat sterol dan memengaruhi aktivitas antimikroba mereka.
Analisis daerah promoter gen SmPR-1 mengungkap banyak elemen cis -regulatori yang terkait dengan respons stres, tahap perkembangan, dan jalur pensinyalan hormonal (Gambar 4 ). Khususnya, situs pengikatan faktor transkripsi MYB dan MYC, yang mengatur pertumbuhan, perkembangan, metabolisme, dan respons tanaman terhadap stresor biotik dan abiotik, lazim ditemukan (Cao et al. 2020 ; Wang et al. 2024 ). Kelimpahan elemen-elemen ini menunjukkan bahwa gen-gen ini kemungkinan terlibat dalam berbagai jalur molekuler dan metabolik. Sebagai gen indikator untuk resistensi yang diperoleh secara sistemik (SAR), famili PR-1 sangat penting untuk pertahanan tanaman terhadap stres biologis (Ali et al. 2018 ), dan akumulasinya meningkatkan resistensi terhadap kondisi yang merugikan (Zribi dan Brini 2020). Dalam penelitian ini, SmPR-1-4 , SmPR-1-7 , dan SmPR-1-9 menunjukkan peningkatan ekspresi secara signifikan pada 12 dan 24 jam pasca infeksi CMV, mempertahankan kadar tinggi selama tahap-tahap selanjutnya. Sebaliknya, SmPR-1-10 dan SmPR-1-11 diekspresikan dengan cepat pada jam ke-6, mencapai puncaknya pada jam ke-12, dan menurun pada jam ke-24. Pola ekspresi yang berbeda ini menunjukkan peran yang bervariasi dari gen PR-1 dalam mekanisme antivirus S. miltiorrhiza , sejalan dengan temuan dalam teh (Zhang et al. 2022a ) dan jelai Tibet (Yin et al. 2023 ). Ekspresi PR-1 yang meningkat telah terbukti meningkatkan resistensi A. thaliana terhadap Peronospora parasitica dan gandum terhadap infeksi karat (Niki et al. 1998 ; Zhang et al. 2015 ). Selain itu, gen seperti TaLr35PR1 , ZmPR-1 , dan ScPR1 memodulasi respons defensif pada gandum, jagung, dan tebu terhadap karat daun (Li et al. 2016 ), Magnaporthe oryzae (Shi 2020 ), dan Acidovorax avenae (Chu et al. 2022 ), masing-masing.
Sistem imun tanaman bergantung pada dua jalur sinyalisasi primer, JA dan SA, yang penting untuk resistensi penyakit melalui interaksi sinergis atau antagonis (Glazebrook 2005 ; Zhang et al. 2010 ). Fitohormon dapat mengaktifkan transkripsi gen PR-1 , yang sangat penting untuk pertahanan tanaman (Pieterse et al. 2009 ). Dalam A. thaliana , ekspresi gen PR-1 diatur oleh SA atau INA (asam 2,6-dikloroisonikotinat) tetapi tidak oleh MeJA atau ET (Durrant dan Dong 2004 ). Sebaliknya, ekspresi PR-1 dalam tembakau memerlukan kombinasi SA, ET, dan MeJA, karena MeJA maupun ET saja tidak cukup (Xu et al. 1994 ). Pada padi, PR-1b diinduksi secara lemah oleh SA tetapi diinduksi secara kuat oleh JA (Mei et al. 2006 ), sedangkan gen BjPR1 pada sawi merespons secara signifikan terhadap SA tetapi tidak terhadap JA atau ABA (Ali et al. 2018 ). Pada kentang, PR-1-5 diinduksi oleh infeksi Phytophthora , ABA, dan IAA (Zaynab et al. 2021 ). Pada jagung, ZmPR-1 diinduksi secara kuat oleh SA, MeJA, dan ABA (Ma et al. 2022 ). Penelitian kami mengungkapkan bahwa famili gen SmPR-1 pada S. miltiorrhiza mengandung elemen respons ABA dan SA dan menunjukkan respons yang bervariasi terhadap ABA, SA, dan BR, yang menunjukkan peran sinergis potensial dalam mekanisme resistensi tanaman.
Penelitian ekstensif telah menunjukkan bahwa gen PR-1 merespons baik stres biotik maupun abiotik, menggarisbawahi peran pentingnya dalam mengelola tantangan lingkungan (Liu et al. 2013 ; Mitsuhara et al. 2008 ; Seo et al. 2008 ; Zeier et al. 2004 ). Selain itu, bukti menunjukkan bahwa protein PR-1 berfungsi di luar respons stres dan berkontribusi pada pertumbuhan dan perkembangan tanaman (Breen et al. 2017 ; Fraser 1981 ; Hanfrey et al. 1996 ). Kehadirannya pada daun pikun tanaman berbunga dewasa dan akumulasinya pada kelopak bunga yang sedang berkembang menunjukkan peran utamanya dalam siklus hidup tanaman (Lotan et al. 1989 ). Pada jagung, 17 gen ZmPR-1 diekspresikan secara berbeda melalui berbagai proses biologis, yang menyoroti keragaman fungsional keluarga gen PR-1 dalam fisiologi tanaman (Ma et al. 2022 ). Dalam penelitian kami, gen SmPR-1 menunjukkan ekspresi yang jauh lebih tinggi pada bunga dibandingkan pada daun S. miltiorrhiza (Gambar 5 ), yang menunjukkan peran pentingnya dalam perkembangan bunga dan pertahanan imun pada spesies ini.
4 Bahan dan Metode
4.1 Identifikasi dan Karakterisasi Gen PR-1
Urutan genom S. miltiorrhiza bersumber dari basis data yang tersedia untuk umum (Ma et al. 2021 ). Untuk mengidentifikasi anggota keluarga gen PR-1 , kami menggunakan basis data Pfam ( http://pfam.xfam.org/ , diakses pada 26 November 2023) untuk analisis (Mistry et al. 2020 ; Sonnhammer et al. 1997 ) untuk mengunduh profil HMM (PF00188), yang merepresentasikan domain terkonservasi yang spesifik untuk keluarga gen ini. Pencarian homologi dilakukan menggunakan alat HMMER dengan ambang batas e -value 1e −10 , diikuti oleh pencarian BLASTP untuk mengonsolidasikan urutan PR-1 yang potensial. Kehadiran domain CAP dalam sekuens yang diidentifikasi diverifikasi menggunakan basis data NCBI CDD ( https://www.ncbi.nlm.nih.gov/cdd/ , diakses pada 27 November 2023) (Marchler-Bauer et al. 2016 ) dan basis data Pfam, dan sekuens yang tidak memiliki domain yang sepenuhnya terkonservasi dikecualikan. SignalP 5.0 ( https://services.healthtech.dtu.dk/service.php?SignalP-5.0 , diakses pada 27 November 2023) digunakan untuk memprediksi peptida sinyal N-terminal dan situs pembelahannya (Armenteros et al. 2019 ). Karakteristik gen tambahan, termasuk titik isoelektrik (pI), berat molekul (BM), grand average hydropathy (GRAVY), dan coding sequence length (CDS), dianalisis menggunakan alat ExPASy ProtParam ( https://web.expasy.org/protparam/ , diakses pada 28 November 2023) (Tabel 1 ) (Gasteiger 2005 ). Lokalisasi subseluler protein SmPR-1 diprediksi menggunakan WoLF PSORT ( https://wolfpsort.hgc.jp/ , diakses pada 28 November 2023) (Armenteros et al. 2019 ).
4.2 Penyelarasan Urutan Komparatif dan Analisis Evolusi
Untuk menyelidiki hubungan evolusi di antara protein PR-1, analisis filogenetik dilakukan menggunakan 88 protein PR-1 dari enam spesies: S. miltiorrhiza (11 protein SmPR-1), Solanum lycopersicum (13 protein SlPR-1), Camellia sinensis (17 protein CsPR-1), Glycine max (24 protein GmPR-1), Arabidopsis thaliana (22 protein AtPR-1), dan Nicotiana tabacum (1 protein NtPR-1). Penyelarasan sekuens berganda dilakukan menggunakan ClustalW dengan pengaturan default, diikuti oleh analisis filogenetik neighbor-joining (NJ) yang dilakukan dalam perangkat lunak MEGA 11 (v11.0.13) untuk membangun pohon filogenetik (Tamura et al. 2021 ). Kekokohan topologi pohon divalidasi menggunakan analisis bootstrap dengan 1000 replikasi.
4.3 Distribusi Kromosom dan Analisis Sintesis Gen Keluarga SmPR-1
Distribusi gen SmPR-1 di seluruh kromosom diperiksa menggunakan data anotasi genom, dengan perancah yang belum dirakit dihapus dari berkas genom. Tekanan selektif pada peristiwa duplikasi gen dinilai dengan menghitung rasio Ka/Ks menggunakan perangkat lunak TBtools. Untuk mengeksplorasi pola duplikasi gen di antara faktor transkripsi SmPR-1 , analisis kolinearitas dilakukan menggunakan MCScanX dan divisualisasikan dengan TBtools. Untuk analisis sinteni, data genom dari beberapa spesies digunakan, termasuk Oryza sativa (beras): PRJDB1747, https://asia.ensembl.org/index.html ; Zea mays (jagung): PRJEB32225, https://www.ncbi.nlm.nih.gov/ ; dan Scutellaria baicalensis (baikal skullcap): GWHBJEC00000000, https://ngdc.cncb.ac.cn/?lang=zh .
4.4 Analisis Struktur Gen, Komposisi Domain Stabil, dan Prediksi Struktur Protein 3D
Fitur arsitektur keluarga gen SmPR-1 dianalisis menggunakan Gene Structure Display Server 2.0 (GSDS) yang tersedia di http://gsds.gao-lab.org/ (diakses pada 30 November 2023). Motif yang dilestarikan diidentifikasi menggunakan alat MEME yang dikonfigurasi untuk mendeteksi hingga 10 motif dengan lebar antara 30 dan 70 asam amino, sementara semua parameter lainnya disimpan pada pengaturan default ( https://meme-suite.org/meme/ , diakses pada 30 November 2023). Prediksi struktur 3D protein dari keluarga gen SmPR-1 dilakukan menggunakan platform SWISS-MODEL ( https://swissmodel.expasy.org/ , diakses pada 23 Januari 2025), dengan pengaturan default.
4.5 Prediksi Elemen Akting Cis Promotor dan Faktor Transkripsi
Untuk mengidentifikasi elemen pengatur cis (CRE) dan pengatur transkripsi gen SmPR-1 , sekuens sepanjang 2000 bp di hulu situs awal transkripsi untuk setiap gen diekstraksi dari data genomik. Basis data PlantCARE ( http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ , diakses pada 04 Desember 2023) digunakan untuk menganalisis daerah promotor CRE (Lescot 2002 ).
4.6 Perawatan Hormonal dan Inokulasi CMV
Daun bibit S. miltiorrhiza berumur 1 bulan dipanen, dan tepi serta tangkai daun dipangkas dan diinokulasi dengan Agrobacterium rhizogenes (strain C58C1) untuk mendorong pertumbuhan akar berbulu. Akar berbulu yang dihasilkan dipindahkan ke labu berbentuk kerucut yang berisi 100 mL larutan nutrisi MS setengah kekuatan dan diinkubasi pada pengocok pada suhu 25°C dan 100 rpm. Setelah sekitar 40 hari, ketika biomassa akar berbulu yang cukup tercapai, kultur diperlakukan dengan larutan BR 50 μM·L −1 . Sampel dikumpulkan pada enam interval waktu: 0,5, 1, 3, 6, 12, dan 24 jam pascaperlakuan. Segera setelah pengumpulan, sampel dibekukan dengan cepat dalam nitrogen cair (−196°C) dan disimpan pada suhu −80°C untuk isolasi RNA. Untuk infeksi CMV, klon cDNA yang menular ditransformasikan ke dalam strain Agrobacterium tumefaciens GV3101 dan diinfiltrasi ke dalam daun S. miltiorrhiza yang baru tumbuh melalui agroinfiltrasi pada OD 600 sebesar 1,0 (Zhou et al. 2021 ). Daun dari tanaman yang terinfeksi CMV dan yang diinokulasi semu dipanen 2 minggu pascainokulasi. Semua percobaan dilakukan dalam rangkap tiga untuk memastikan replikasi biologis.
4.7 Isolasi RNA dan Persiapan cDNA
Total RNA diekstraksi dari sampel menggunakan metode penggilingan dan dimurnikan menggunakan RNA Easy Fast Plant Tissue Total RNA Isolation Kit (TIANGEN, Tiongkok). RNA yang dimurnikan mengalami transkripsi balik menggunakan Evo M-MLV RT Mix Kit (Accurate Biology, Tiongkok) untuk mensintesis DNA komplementer (cDNA). cDNA yang dihasilkan disimpan pada suhu -80°C untuk analisis selanjutnya.
4.8 Deteksi RNA-Seq dan qRT-PCR
Data transkriptom untuk jaringan S. miltiorrhiza diperoleh dari laboratorium kami (saat ini belum dipublikasikan), dan data transkriptom tambahan untuk sampel yang diobati dengan ABA, MeJA, dan SA diperoleh dari basis data BioProject menggunakan pengenal PRJNA703309 (ABA), PRJNA393563 (MeJA), dan PRJNA301529 (SA) ( https://www.ncbi.nlm.nih.gov/ , diakses pada 16 Desember 2024). Untuk menyusun peta regulasi famili SmPR-1 , urutan protein famili SmPR-1 digunakan sebagai urutan kueri, dan fungsi BLAST dalam TBtools (v2.154) digunakan untuk mencari urutan protein famili SmPR-1 yang cocok dalam pustaka protein transkriptom. Nilai FPKM dari urutan yang cocok kemudian diekstraksi. Untuk menyajikan tren regulasi secara lebih intuitif, nilai FPKM untuk titik waktu berbeda dari perlakuan yang sama dinormalisasi menggunakan fungsi distribusi normal (Z = (X—μ)/σ, di mana X adalah nilai FPKM, μ adalah mean, dan σ adalah simpangan baku), dan peta panas ekspresi divisualisasikan menggunakan TBtools (v2.154) (Chen et al. 2020 ). Urutan primer untuk gen SmPR-1 dirancang menggunakan Real-time PCR (TaqMan) dan Probes Design Tool ( https://www.genscript.com/tools/real-time-pcr-taqman-primer-design-tool , diakses pada 13 Desember 2023) (Tabel S1 ). Tingkat ekspresi gen SmPR-1 diukur melalui qRT-PCR menggunakan cDNA yang disintesis dari jaringan bunga sebagai templat dan aktin sebagai kontrol internal. Setiap reaksi mengandung 1 μL cDNA cetakan, 0,5 μL masing-masing primer hulu dan hilir, 5 μL 2X SYBR Green Pro Taq HS Premix (Accurate Biology, Cina), dan 3 μL ddH 2 O untuk total volume reaksi 10 μL. Protokol qRT-PCR mencakup denaturasi awal pada 95 °C selama 30 detik, diikuti oleh 40 siklus 5 detik pada 95 °C dan 30 detik pada 60 °C, dengan pemantauan fluoresensi. Analisis kurva leleh dilakukan dengan langkah-langkah pada 95 °C selama 15 detik, 60 °C selama 1 menit, dan 95 °C selama 1 detik, dengan fluoresensi direkam pada setiap langkah. Ekspresi gen diukur dengan metode 2 -ΔΔCt . Setiap sampel diukur dalam rangkap tiga untuk memastikan keandalan hasil.
4.9 Kloning Gen dan Analisis Lokalisasi Subseluler
Urutan pengkodean (CDS) SmPR-1-5 dikloning ke dalam vektor ekspresi transien pHB (Tabel S1 ), yang berisi penanda fluoresensi kuning. Pengujian agroinfiltrasi dilakukan menurut protokol standar (Shi et al. 2022 ). Daun dari tanaman N. benthamiana berumur 1 bulan dipilih untuk infiltrasi, dan suspensi bakteri diinokulasi ke epidermis daun bagian bawah dan ditandai dengan tepat. Setelah infiltrasi, tanaman dipelihara dalam kondisi optimal, termasuk inkubasi gelap selama 24 jam pada suhu 25°C, diikuti oleh inkubasi terang selama 24 jam. Vektor kosong yang disiapkan dalam kondisi identik berfungsi sebagai kontrol. Lokalisasi subseluler protein SmPR-1-5 diperiksa menggunakan mikroskop fluoresensi konfokal laser, dan emisi fluoresensi hijau direkam menggunakan mikroskop konfokal pemindaian laser Zeiss LSM 880. Untuk mengonfirmasi hasil ini, alat daring WoLF PSORT ( http://wolfpsort.seq.cbrc.jp/ , diakses pada 5 Maret 2024) digunakan untuk memprediksi lokalisasi subseluler protein yang dikodekan SmPR-1 .
5 Kesimpulan
Studi ini melakukan investigasi genomik komprehensif dari keluarga gen PR-1 di S. miltiorrhiza dan mengidentifikasi 11 gen SmPR-1 melalui analisis bioinformatika. Gen-gen ini mengkode protein dengan domain CAP yang terkonservasi dan peptida sinyal. Analisis filogenetik mengelompokkan gen SmPR-1 ke dalam lima klade berbeda, yang menunjukkan divergensi evolusioner dan spesialisasi fungsional potensial. Analisis lokalisasi kromosom menunjukkan bahwa keluarga gen SmPR-1 didistribusikan pada Chr1 (6 gen), Chr6 (3 gen), dan Chr8 (1 gen). Analisis kolinearitas intraspesifik mengidentifikasi duplikasi segmental SmPR-1-5 dan SmPR-1-11 pada Chr1. Analisis interspesifik menunjukkan bahwa lima gen SmPR-1 kolinear dengan A. thaliana dan S. baicalensis . Profil ekspresi spesifik jaringan mengungkapkan tingkat ekspresi tinggi gen SmPR-1 tertentu pada bunga dan batang, yang menunjukkan perannya dalam proses perkembangan. Studi lokalisasi subseluler menunjukkan bahwa SmPR-1-5 hadir di sitoplasma dan nukleus, yang menyiratkan keterlibatannya dalam jalur pensinyalan intraseluler. Analisis ekspresi di bawah tekanan biotik dan abiotik menyoroti regulasi hormonal yang signifikan dari gen SmPR-1 , dan data transkriptom dari tanaman yang terinfeksi CMV mengonfirmasi responsivitasnya terhadap stres biologis, yang menggarisbawahi perannya dalam mekanisme pertahanan tanaman. Temuan ini memberikan wawasan berharga tentang fungsi molekuler gen SmPR-1 dalam respons stres dan resistensi penyakit. Studi ini membangun landasan untuk studi masa depan yang mengeksplorasi peran gen PR-1 dalam imunitas tanaman dan pengembangan kultivar S. miltiorrhiza dengan peningkatan resistensi penyakit. Validasi eksperimental lebih lanjut dan eksplorasi interaksi gen PR-1 dengan jalur pertahanan lainnya sangat penting untuk sepenuhnya memahami kontribusinya terhadap mekanisme pertahanan tanaman.

