
ABSTRAK
DNA sirkular ekstrakromosomal (eccDNA) memainkan peran penting dalam mengatur berbagai proses biologis, termasuk perkembangan otot yang abnormal. Fungsi molekuler dan dampak eccDNA dalam perkembangan otot ikan kurang dipahami. Untuk menyelidiki peran potensial eccDNA dalam perkembangan otot ikan, kami menganalisis dan membandingkan profil ekspresi eccDNA otot ikan mas rumput renyah, yang diberi diet berbasis tepung kacang faba, dan ikan mas rumput biasa, yang diberi diet praktis. Dengan menggunakan strategi Circle-seq, kami menemukan kelimpahan eccDNA pada ikan mas rumput renyah (211.920 eccDNA) secara signifikan lebih tinggi daripada yang ada pada ikan mas rumput biasa (25.857 eccDNA), yang menunjukkan bahwa diet kacang faba kemungkinan secara independen memengaruhi produksi eccDNA. Dibandingkan dengan ikan mas rumput biasa, ikan mas rumput renyah menunjukkan 10.565 eccDNA yang diatur naik dan 129 eccDNA yang diatur turun, yang menunjukkan eccDNA mungkin terkait dengan perkembangan otot ikan mas rumput. Analisis pengayaan GO dan KEGG menunjukkan bahwa eccDNA yang meningkat terkait dengan perkembangan serat otot, struktur seluler, dan sambungan sel. Berdasarkan hasil kami, kami berspekulasi bahwa ekspresi berlebihan gen yang terlibat dalam serat otot, metabolisme kalsium, dan kolagen yang didorong oleh eccDNA kemungkinan berkontribusi pada peningkatan yang diamati dalam kepadatan serat otot, kadar kalsium, dan kandungan kolagen pada ikan mas rumput renyah, sehingga meningkatkan kekerasan otot. Khususnya, eccDNA diidentifikasi sebagai imunostimulan bawaan potensial yang mampu menimbulkan respons imun pada ikan. Singkatnya, temuan kami menunjukkan bahwa eccDNA diekspresikan secara tidak normal dalam otot ikan yang diberi makan kacang faba, menawarkan wawasan baru tentang mekanisme molekuler yang mendasari pengerasan otot pada ikan.
1 Pendahuluan
Tekstur otot merupakan penentu penting kualitas ikan, yang secara langsung memengaruhi produksi produk ikan berkualitas tinggi (Hyldig dan Nielsen 2001 ). Memahami mekanisme molekuler yang mengatur tekstur otot ikan dapat memajukan pengembangan produk ikan premium secara signifikan dan meningkatkan keuntungan dalam industri akuakultur (Elvevoll et al. 1996 ). Di antara karakteristik otot, kekerasan, atau kekencangan, sangat penting dalam membentuk preferensi konsumen (Veland dan Torrissen 1999 ). Ikan mas rumput ( Ctenopharyngodon idellus ), ikan air tawar asli Tiongkok, memiliki kepentingan ekonomi yang signifikan (Xu et al. 2020 ), dengan produksi melebihi 5,7 juta ton pada tahun 2022, yang mencakup seperlima dari hasil akuakultur air tawar Tiongkok. Percobaan penting menunjukkan bahwa memberi makan ikan mas rumput dengan kacang faba ( Vicia faba ) selama 90–120 hari secara signifikan meningkatkan kerenyahan dan kekencangan tekstur otot mereka (Yu et al. 2017 ; Hao et al. 2024 ). Hal ini menyebabkan peningkatan popularitas ikan mas rumput renyah di Tiongkok, dengan produk-produknya berhasil menembus pasar di Amerika Serikat, Asia Tenggara, dan Amerika Latin. Selain itu, diameter dan kepadatan serat otot sangat berkorelasi dengan kekerasan otot (Johnston et al. 2000 ; Yu et al. 2017 ). Dibandingkan dengan ikan mas rumput biasa, ikan mas rumput renyah menunjukkan diameter serat otot yang berkurang dan kepadatan serat yang meningkat (Yu, Xie et al. 2014 ; Feng et al. 2016 ; Tang et al. 2024 ). Menariknya, sementara komposisi otot langsungnya (lipid, air, atau protein) tidak menunjukkan perbedaan yang signifikan (Tian et al. 2020 ; Hao et al. 2024 ; Ma et al. 2024 ), ikan mas rumput renyah menunjukkan kandungan kolagen yang lebih tinggi dan serat otot yang lebih kecil (Ma et al. 2020 ). Meskipun penelitian sebelumnya mengidentifikasi beberapa protein dan gen yang berpotensi terkait dengan kekencangan otot, mekanisme molekuler yang mendorong peningkatan kekerasan otot pada ikan mas rumput renyah masih kurang dipahami (Yu et al. 2020 ; Fu et al. 2022 ; Tian et al. 2023 ).
DNA sirkular ekstrakromosom (eccDNA) adalah molekul DNA unik yang dicirikan oleh arsitektur untai ganda dan sirkular. DNA ini berasal dari DNA genomik dan ada secara independen dari DNA kromosom (Wang et al. 2021 ). Berkisar dalam ukuran dari ratusan basa hingga megabasa, eccDNA lazim ditemukan pada eukariota, termasuk ragi, nematoda, lalat buah, tumbuhan, dan mamalia (Møller et al. 2016 ; Kumar et al. 2017 ). EccDNA dapat bereplikasi secara otonom dan menyimpang dari pewarisan Mendel, dengan distribusinya yang tidak merata pada sel anak meningkatkan efisiensi transkripsi gen dibandingkan dengan DNA kromosom (Wu et al. 2019 ; Lv et al. 2024 ). Mereka memiliki kapasitas untuk memengaruhi fenotipe dengan menyesuaikan kuantitas salinan gen dan transkripsi gen panjang penuh atau terpotong (Nathanson et al. 2014 ; Paulsen et al. 2018 ; Zhu et al. 2021 ). Selain itu, eccDNA yang mengandung elemen pengatur, seperti enhancer, dapat mengatur amplifikasi gen melalui interaksi intramolekuler (Sheng et al. 2024 ). Bukti yang muncul menunjukkan bahwa eccDNA dikaitkan dengan pengaturan perkembangan otot. Misalnya, kuantitas tertinggi eccDNA telah ditemukan pada gen pengkode protein otot manusia titin (TTN), yang menunjukkan bahwa eccDNA dapat membantu sel inang dalam menjalankan peran yang dimaksudkan (Møller et al. 2018 ). Demikian pula, eccDNA yang berasal dari gen AGRIN, yang diperkaya dalam otot burung dara raja, mengkode protein membran yang penting untuk perkembangan sambungan neuromuskular, dengan mutasi yang menyebabkan perkembangan otot yang abnormal (Møller et al. 2020 ). Baru-baru ini, kami mengungkapkan bahwa eccDNA yang terlibat dalam karakteristik otot sangat melimpah dalam sel otot ikan mas rumput yang sedang melangsingkan dibandingkan dengan ikan mas rumput biasa, yang menunjukkan bahwa eccDNA yang diperkaya mungkin memiliki efek pada aktivasi kekencangan otot (He et al. 2024 ).
Meskipun demikian, peran eccDNA dalam perkembangan otot ikan masih belum jelas. Penelitian ini bertujuan untuk meneliti eccDNA yang ada dalam otot ikan mas rumput yang diberi kacang faba, dan meneliti perannya dalam mengatur tekstur otot ikan. Temuan ini memberikan dasar untuk meningkatkan kualitas fillet ikan dalam akuakultur.
2 Bahan dan Metode
2.1 Pengumpulan Sampel dan Ekstraksi DNA
Sebanyak 120 ikan dialokasikan secara acak ke dalam kelompok ikan mas rumput biasa dan ikan mas rumput renyah, dengan tiga kali ulangan per kelompok perlakuan. Mereka dibesarkan dalam enam tangki (20 ikan di setiap tangki) di sebuah peternakan air di Zhongshan, Guangdong, Cina. Ikan mas rumput yang diberi diet praktis disebut sebagai ikan mas rumput biasa, sedangkan yang diberi makan secara eksklusif dengan tepung kacang faba ( Vicia faba ) disebut ikan mas rumput renyah. Komposisi diet praktis konsisten dengan yang digunakan dalam penelitian oleh Gan et al. ( 2017 ). Kedua kelompok dibesarkan dalam kondisi akuakultur konvensional (suhu air 25°C, pH = 7,0, dan oksigen terlarut 5 mg/L). Berat akhir sekitar 4,5 kg untuk kelompok ikan mas rumput renyah dan sekitar 5 kg untuk kelompok ikan mas rumput biasa setelah tiga bulan. Tekstur otot ikan mas rumput biasa relatif lembut, sedangkan tekstur otot ikan mas rumput renyah secara signifikan lebih kencang dan keras, sehingga keduanya dapat dibedakan. Tiga ekor ikan mas rumput biasa dan tiga ekor ikan mas rumput renyah dipilih secara acak dan dieutanasia dengan cara direndam dalam eugenol (80 mg/L). Sekitar 10 g jaringan otot punggung diambil dari setiap sampel dan segera dikriopreservasi dalam nitrogen cair. Ekstraksi DNA genom dilakukan menggunakan MagAttract HMW DNA Kit (QIAGEN), sesuai dengan petunjuk pabrik pembuatnya. Kami menilai kualitas DNA yang diekstraksi melalui elektroforesis pada gel agarosa yang diwarnai etidium bromida 1%. Konsentrasi dan kemurnian DNA diukur menggunakan NanoDrop Microvolume Spectrophotometer (Thermo Scientific).
2.2 Penghapusan DNA Linier
Untuk memperkaya DNA sirkular, sisa DNA genomik linear dihilangkan dari setiap sampel. DNA dicerna pada suhu 37°C selama 120 jam menggunakan Plasmid-Safe ATP-dependent DNase (Biosearch, E3110K), mengikuti protokol pabrik. DNase dan ATP tambahan ditambahkan setiap 24 jam. Efektivitas penghilangan DNA linear diverifikasi melalui amplifikasi PCR dari gen penanda DNA sirkular ( COX3 ) dan gen penanda DNA linear ( NDL6 ) Amplifikasi PCR dilakukan dengan denaturasi awal pada suhu 94°C selama 5 menit, diikuti oleh 35 siklus denaturasi pada suhu 94°C selama 30 detik, annealing pada suhu 55°C ( COX3 ) atau 62°C ( NDL6 ) selama 30 detik, dan ekstensi pada suhu 72°C selama 60 detik. Ekstensi akhir dilakukan pada suhu 72°C selama 5 menit. Produk PCR dievaluasi melalui elektroforesis gel agarosa. Kehadiran pita COX3 spesifik dan tidak adanya pita NDL6 mengonfirmasi penghapusan lengkap DNA genom linier.
2.3 Pengayaan DNA Sirkular
Setelah penghilangan DNA linear, DNA sirkuler diamplifikasi secara acak menggunakan polimerase Phi29 dan primer acak yang tahan terhadap eksonuklease melalui amplifikasi lingkaran bergulir (RCA). Reaksi RCA dilakukan pada suhu 30°C selama 72 jam. EccDNA yang diamplifikasi dimurnikan menggunakan Cycle-Pure Kit (Omega) dan dicerna dengan NdeI (Thermo Fisher Scientific) untuk memverifikasi hasil RCA sebelum pengurutan.
2.4 Identifikasi DNA Sirkular
Program Circle-Map (Prada-Luengo et al. 2019 ) digunakan untuk mengidentifikasi eccDNA. Untuk meningkatkan akurasi, parameter penyaringan meliputi: “(1) Skor lingkaran >=200, (2) Pemetaan pembacaan terpisah >= 2, (3) Peningkatan cakupan situs awal >= 0,33, (4) Peningkatan cakupan koordinat terminal >= 0,33, (5) Keseragaman cakupan sekuensing <= 0,1, dan (6) Cakupan rata-rata > Simpangan baku”. Untuk pemeriksaan lebih lanjut, semua eccDNA yang memenuhi persyaratan penyaringan digunakan.
2.5 Wilayah Sumber dan Anotasi EccDNA
Untuk menganalisis distribusi genomik eccDNA, setiap kromosom dibagi menjadi jendela 50 kb, dan kelimpahan eccDNA dalam setiap jendela dihitung. Plot Manhattan dibuat untuk merepresentasikan distribusi secara visual. Urutan gen pengkode protein dari genom referensi ikan mas rumput digunakan (Wu et al. 2022 ). BEDtools digunakan untuk mengidentifikasi tumpang tindih antara urutan gen yang dianotasi dan koordinat eccDNA (Quinlan 2014 ). Skrip R khusus digunakan untuk menyaring gen eccDNA yang dianotasi, mempertahankan gen dengan jumlah pasangan basa yang tumpang tindih melebihi 60 untuk analisis lebih lanjut.
2.6 Analisis Diferensial Ekspresi DNA Sirkular
Data jumlah tag eccDNA dianalisis menggunakan perangkat lunak edgeR untuk analisis ekspresi diferensial (Robinson et al. 2010 ). Analisis tersebut mencakup tiga langkah utama: (1) normalisasi jumlah tag; (2) perhitungan probabilitas pengujian hipotesis ( nilai- p ), tergantung pada model; dan (3) pengujian hipotesis berganda dan koreksi untuk menentukan nilai FDR (tingkat deteksi kesalahan). eccDNA dengan nilai- p < 0,05 dan |log 2 FC| > 1 diidentifikasi sebagai diekspresikan secara diferensial berdasarkan analisis.
2.7 Analisis Pengayaan Fungsional dan Jalur Gen Terkait DNA Sirkular
Gen terkait eccDNA yang diekspresikan secara berbeda menjadi sasaran analisis pengayaan fungsional menggunakan Gene Set Enrichment Analysis (GSEA, http://www.gseamsigdb.org/gsea/msigdb/annotate.jsp ). Analisis pengayaan jalur Kyoto Encyclopedia of Genes and Genomes (KEGG) dan Gene Ontology (GO) dilakukan untuk menjelaskan fungsi dan jalur biologis. Visualisasi eccDNA dilakukan menggunakan perangkat lunak IGV (Versi 2.4.10) (Thorvaldsdottir et al. 2013 ).
2.8 Analisis Statistik
Analisis statistik dilakukan menggunakan R-4.1.2. Untuk penyaringan eccDNA yang terwakili secara berlebihan untuk ikan kerapu dari eccDNA yang diberi anotasi, signifikansi eccDNA dalam dua kelompok dianalisis melalui R-4.1.2 dan dinilai melalui nilai p uji Wilcoxon rank-sum . Hasil dianggap signifikan secara statistik jika nilai p kurang dari 0,05.
3 Hasil
3.1 Deteksi Penghapusan DNA Linier dan Pengayaan DNA Sirkular
Hasil elektroforesis yang menilai amplifikasi gen COX3 dan NDL6 , yang digunakan untuk mengevaluasi penghilangan DNA genomik linear, ditunjukkan pada Gambar 1A,B , berturut-turut. Pita yang jelas diamati untuk COX3 , sementara tidak ada pita yang terdeteksi untuk NDL6 , yang menunjukkan penghilangan DNA genomik linear secara menyeluruh.
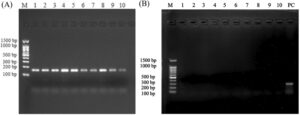
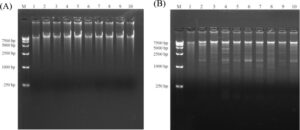
Gambar 2A,B menggambarkan hasil elektroforesis setelah reaksi RCA dan pencernaan enzim berikutnya untuk pemurnian eccDNA. Reaksi RCA berhasil mengonsentrasikan eccDNA utuh, yang setelah pencernaan enzimatik menghasilkan fragmen dengan berbagai ukuran.
3.2 Identifikasi dan Karakteristik Genom eccDNA
Pembacaan sekuensing mentah dibuat menggunakan platform Illumina NovaSeq 6000. Data diproses menggunakan Trimmomatic untuk menghilangkan pembacaan berkualitas rendah (yang berisi sepuluh atau lebih basa yang tidak diurutkan atau berkualitas rendah), duplikat PCR, dan sekuens adaptor. Jumlah pembacaan bersih yang diperoleh dari enam sampel (ikan mas rumput renyah: 1–3; ikan mas rumput biasa: 1–3) adalah sebagai berikut: 119.020.284 bp, 147.929.354 bp, 106.936.632 bp, 139.871.412 bp, 123.377.174 bp, dan 130.942.792 bp, masing-masing (Tabel 1 ). Deteksi EccDNA dilakukan menggunakan pendekatan Circle-Seq pada tiga sampel dari masing-masing kelompok ikan mas rumput renyah dan biasa. Secara total, 237.777 eccDNA teridentifikasi, dengan 211.920 berasal dari ikan kerapu dan 25.857 berasal dari ikan kerapu biasa. Hasil ini menunjukkan bahwa eccDNA ada di mana-mana dalam otot ikan dan secara signifikan lebih banyak dalam ikan kerapu.
| Nama sampel | Bacaan mentah | Bacaan bersih | Jumlah 30 (%) |
|---|---|---|---|
| Ikan mas rumput renyah 1 | 135.401.198 | 119.020.284 | 90.75 |
| Ikan mas rumput renyah 2 | 169.148.422 orang | 147.929.354 orang | 90.46 |
| Ikan mas rumput renyah 3 | 121.513.794 orang | 106.936.632 orang | 89.95 |
| Ikan mas rumput biasa 1 | 157.244.712 orang | 139.871.412 | 91.78 |
| Ikan mas rumput biasa 2 | 139.295.000 | 123.377.174 orang | 91.32 |
| Ikan mas rumput biasa 3 | 153.561.782 orang | 130.942.792 orang | 91.82 |
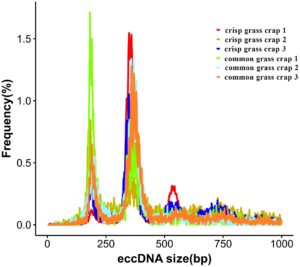
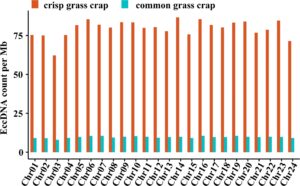
Analisis mengungkapkan bahwa panjang eccDNA pada otot ikan mas rumput sangat bervariasi, terutama berkisar antara 100 hingga 600 bp, dengan dua puncak pada sekitar 185 bp dan 300 bp (Gambar 3 ). Distribusi ukuran ini sejalan dengan temuan dari penelitian sebelumnya pada eccDNA otot (Møller et al. 2020 ). Urutan eccDNA pada otot ikan mas rumput didistribusikan di seluruh kromosom. Analisis frekuensi eccDNA per megabasa (Mb) pada setiap kromosom menunjukkan bahwa sebagian besar kromosom menunjukkan frekuensi yang melebihi 75 eccDNA per Mb. Khususnya, kromosom 14, 16, dan 23 menunjukkan frekuensi yang relatif lebih tinggi dibandingkan dengan yang lain (Gambar 4 ).
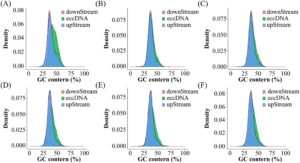
Gambar 5A–Fd mengilustrasikan analisis kandungan GC dalam wilayah eccDNA dan wilayah pengapitnya sepanjang 1000 bp di hulu dan hilir. Kandungan GC dalam wilayah ini berkisar antara 25% hingga 60%, dengan puncak sekitar 35%.
3.3 Profil Ekspresi Diferensial dan Anotasi eccDNA
Untuk menyelidiki dasar molekuler kekencangan fillet ikan, analisis komparatif profil ekspresi eccDNA antara jaringan otot ikan mas rumput renyah dan biasa dilakukan. Sebanyak 10.694 eccDNA menunjukkan ekspresi diferensial antara kedua kelompok. Ikan mas rumput renyah menunjukkan 10.565 eccDNA yang teregulasi naik dan 129 eccDNA yang teregulasi turun dibandingkan dengan ikan mas rumput biasa. Analisis anotasi mengungkapkan bahwa eccDNA yang teregulasi naik dikaitkan dengan 22.186 gen, sedangkan eccDNA yang teregulasi turun dikaitkan dengan 59 gen.
3.4 Analisis Pengayaan GO dan KEGG pada eccDNA yang Diekspresikan Secara Berbeda
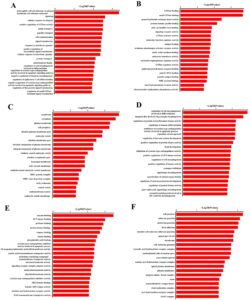
Analisis pengayaan GO menunjukkan bahwa gen yang berhubungan dengan eccDNA yang teregulasi naik berhubungan dengan komponen seluler (aktivitas faktor pertukaran guanyl-nukleotida, pengikatan GTPase, dan pengikatan GTPase kecil dan), proses biologis (respons seluler dan pensinyalan terhadap stimulus) dan fungsi molekuler (membran, sitoplasma dan membran plasma) (Gambar 6A–C ). Untuk eccDNA yang teregulasi turun, gen terkait diperkaya dalam proses biologis (pengaturan morfogenesis sel selama diferensiasi, morfogenesis tepi sayap yang berasal dari cakram imajiner dan pengaturan aktivitas protein serin/treonin kinase), komponen seluler (pengikatan kinase JUN, pengikatan protease dan pengikatan enzim) dan fungsi molekuler (sambungan sel, sambungan adheren dan sambungan penjangkaran) (Gambar 6D,E ).
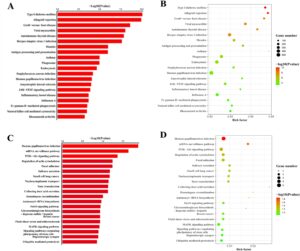
Lebih jauh, analisis pengayaan jalur KEGG mengidentifikasi 383 jalur yang mengalami peningkatan regulasi yang terkait dengan gen terkait eccDNA yang diekspresikan secara berbeda. Jalur-jalur ini meliputi jalur seperti penolakan allograft, diabetes melitus tipe I, miokarditis virus, penyakit graft-versus-host, infeksi virus herpes simpleks 1, penyakit tiroid autoimun, campak, pemrosesan dan penyajian antigen, asma, dan jalur pensinyalan JAK–STAT (Gambar 7A–B ). Sebaliknya, 96 jalur yang mengalami penurunan regulasi diidentifikasi, meliputi infeksi human papillomavirus, jalur pengawasan mRNA, dan jalur pensinyalan PI3K-Akt (Gambar 7C,D ).
3.5 Analisis Gen eccDNA yang Diekspresikan Secara Berbeda Terkait dengan Kekerasan Otot
Analisis lebih lanjut difokuskan pada gen-gen yang diekspresikan secara berbeda yang terkait dengan kekerasan otot pada ikan mas rumput. Ikan mas rumput crisp menunjukkan eccDNA yang meningkat yang sesuai dengan gen-gen yang terlibat dalam struktur serat otot, perkembangan otot, organisasi sitoskeletal, dan metabolisme kalsium. Ini termasuk gen-gen yang mengkode kolagen, talin, catenin, tubulin, dan protein pengikat miosin-C (Tabel 2 ). Kandidat eccDNA ini kemungkinan berkontribusi pada peningkatan kekerasan otot yang diamati pada ikan mas rumput crisp.
| Deskripsi gen | Anotasi | Peraturan | catatan 2 FC | P |
|---|---|---|---|---|
| Struktur dan perkembangan serat otot | ||||
| Rantai berat miosin | GC01Gen03301 | Ke atas | 20.87947345 | 0,000000093 |
| Rantai ringan miosin | GC01Gen01667 | Ke atas | nomor telepon 20.84801687 | 0,0000000972 |
| Aktin, alfa | GC01Gen01404 | Ke atas | nomor telepon 21.40380748 | 0,0000000436 |
| Protein terkait aktin | GC01Gen01316 | Ke atas | Nomor telepon 10.0231253 | 0,010311326 |
| Protein pengikat miosin | GC01Gen27142 | Ke atas | nomor telepon 20.95861459 | 0,000000083 |
| Protein pengikat miosin C, tipe a cepat | GC01Gen04318 | Ke atas | 10.32899188 | 0,008203815 |
| Troponin C tipe 1b (lambat) | GC01Gen27309 | Ke atas | 8.33492794 | 0,032969156 |
| Tropomiosin | GC01Gen03047 | Ke atas | nomor telepon 20.84801687 | 0,0000000972 |
| Pengulangan Nebulin | GC01Gen02061 | Ke atas | nomor telepon 21.47093962 | 0,0000000396 |
| Sitoskeleton | ||||
| Tabung | GC01Gen01232 | Ke atas | nomor telepon 21.40380748 | 0,0000000436 |
| Kata “Katenin” | GC01Gen01138 | Ke atas | nomor telepon 21.40380748 | 0,0000000436 |
| Klatrin | GC01Gen14416 | Ke atas | 8.522483704 | 0,029215294 |
| Fosfolipase C | GC01Gen02070 | Ke atas | nomor telepon 21.47093962 | 0,0000000396 |
| Pembatasan protein | GC01Gen07223 | Ke atas | Nomor telepon 9.169432211 | 0,018953284 |
| Nexilin (protein pengikat F Actin) | GC01Gen11576 | Ke atas | Nomor telepon 21.12714634 | 0,0000000652 |
| Talin, domain tengah | GC01Gen13969 | Ke atas | nomor telepon 20.84801687 | 0,0000000972 |
| Talin 2a | GC01Gen29803 | Ke atas | 6.80371429 | 0,025924749 |
| Kolagen | ||||
| Kolagen tipe I alfa | GC01Gen04089 | Ke atas | 10.32899188 | 0,008203815 |
| Kolagen tipe II, alfa-1a | GC01Gen11512 | Ke atas | Nomor telepon 21.12714634 | 0,0000000652 |
| Metabolisme kalsium | ||||
| Kalretikulin | GC01Gen03052 | Ke atas | nomor telepon 20.84801687 | 0,0000000972 |
| Kalmodulin | GC01Gen16386 | Ke atas | 8.745439622 | 0,025237536 |
| Protein terkait kadherin | GC01Gen01138 | Ke atas | nomor telepon 21.40380748 | 0,0000000436 |
| Lampiran | GC01Gen00727 | Ke atas | Nomor telepon 21.12714634 | 0,0000000652 |
| Protokaderin 10 | GC01Gen00569 | Ke atas | Nomor telepon 21.12714634 | 0,0000000652 |
| Berbentuk belah ketupat, seperti urat 3 | GC01Gen08395 | Ke atas | nomor telepon 8.059328516 | 0,039232168 |
4 Diskusi
Tekstur otot merupakan penentu utama kualitas produk ikan, yang secara langsung memengaruhi preferensi konsumen dan nilai pasar. Pemahaman yang lebih mendalam tentang mekanisme biologis yang mendasari tekstur otot ikan dapat memfasilitasi pengembangan produk bernilai tinggi dan meningkatkan profitabilitas dalam industri akuakultur. Ikan mas rumput renyah, yang diproduksi dengan memberi makan ikan mas rumput secara eksklusif dengan kacang faba, menunjukkan tekstur otot yang lebih unggul dibandingkan dengan ikan mas rumput biasa. Kualitas tekstur yang ditingkatkan ini mencakup peningkatan kekenyalan, kekencangan, kekenyalan, gaya geser, dan kekenyalan, menjadikan ikan mas rumput renyah sebagai produk akuakultur premium (Chen et al. 2020 ). Penelitian sebelumnya terutama difokuskan pada pemahaman mekanisme yang meningkatkan tekstur otot pada ikan mas rumput, dengan tujuan menghasilkan fillet ikan berkualitas tinggi. Penelitian ini merupakan kemajuan yang signifikan, karena merupakan yang pertama membandingkan profil eccDNA antara jaringan otot ikan mas rumput renyah dan biasa, yang memberikan wawasan berharga untuk penelitian masa depan tentang kekencangan fillet ikan.
ROS seperti hidrogen peroksida (H 2 O 2 ) diproduksi oleh dua turunan aminopirimidin glukosidik (vicine dan convicine) yang banyak terdapat dalam kacang faba (Winterbourn et al. 1986 ). Studi metabolomik melaporkan peningkatan kadar ROS secara signifikan (hingga empat kali lipat) pada otot rangka ikan mas rumput renyah (Yu et al. 2020 ). ROS diketahui memainkan peran penting dalam mengatur metabolisme otot dan cenderung memengaruhi tekstur otot (Archile-Contreras dan Purslow 2011 ). Namun, sementara kacang faba meningkatkan tekstur otot, mereka juga menghambat pertumbuhan dan dapat menyebabkan kekurangan nutrisi dan kerusakan oksidatif (Ma et al. 2020 ; Fu et al. 2022 ). Dalam studi ini, kami mengamati bahwa kelimpahan eccDNA dalam otot ikan mas rumput renyah (211.920) secara signifikan lebih tinggi daripada pada otot ikan mas rumput biasa (25.857). Selain itu, 10.823 eccDNA yang diekspresikan secara diferensial diidentifikasi antara otot ikan mas rumput renyah dan otot ikan mas rumput biasa, termasuk 10.694 eccDNA yang diatur naik dan 129 yang diatur turun pada otot ikan mas rumput renyah. Peningkatan stres oksidatif telah terbukti memperburuk kerusakan kromosom, yang mengarah pada kerusakan untai ganda (DSB) dan kerusakan untai tunggal (SSB), serta meningkatkan oksidasi basa (Dutta et al. 2015 ; Cannan dan Pederson 2016 ). Proses-proses ini memfasilitasi pembentukan molekul DNA melingkar, yang dapat menjelaskan kelimpahan eccDNA yang lebih tinggi yang diamati pada ikan mas rumput renyah. Kami mengusulkan bahwa peningkatan stres oksidatif yang disebabkan oleh pemberian kacang faba memfasilitasi pelepasan DNA dari kromosom dan sirkularisasi berikutnya dalam otot ikan.
Kualitas daging ikan dan massa otot sangat dipengaruhi oleh dua faktor penting: ukuran dan jumlah serat otot (Tang et al. 2024 ). Temuan sebelumnya telah menunjukkan bahwa serat otot terkait erat dengan peningkatan kekerasan otot ikan, dan pertumbuhan otot yang diinduksi kacang faba pada ikan mas rumput merupakan hasil dari hiperplasia, yang ditandai dengan peningkatan jumlah miofiber seumur hidup (Johnston et al. 2000 ). Miofibril, unit fungsional serat otot, terutama terdiri dari filamen tipis (terdiri dari protein troponin, aktin, dan tropomiosin), filamen tebal (mengandung protein miosin), titin, garis-M, dan cakram-Z (terdiri dari protein nebulin dan desmin) (Nishimura 2010 ). Protein miofibril merupakan kelas protein dominan dalam otot rangka, meliputi sekitar 60% dari total komposisi protein berdasarkan berat (Huang et al. 2012 ). Temuan sebelumnya menunjukkan bahwa gen yang terkait dengan proliferasi miofibroblast dan produksi sitokin meningkat pada tingkat mRNA dan tingkat protein pada ikan mas rumput renyah dibandingkan dengan ikan mas rumput biasa (Yu, Xie et al. 2014 ; Yu, Liu et al. 2014 ; Yu et al. 2017 ; Xu et al. 2020 ). Secara kebetulan, dalam penelitian saat ini, analisis pengayaan fungsional dari eccDNA yang meningkat pada ikan mas rumput renyah mengungkapkan asosiasi signifikan dengan gen yang terkait dengan struktur serat otot, perkembangan sel otot, dan diferensiasi. Ini termasuk gen yang mengkode protein capping, miosin, tropomiosin, talin, annexin, catenin, clathrin, tubulin, dan lainnya. Temuan ini menunjukkan bahwa regulasi molekuler pengerasan otot secara substansial dipengaruhi oleh eccDNA yang meningkat yang diperkaya dalam fungsi terkait otot. Peningkatan aktivitas gen kromosom dan ekstrakromosom, jumlah salinan yang tinggi, dan peningkatan aktivitas transkripsi eccDNA kemungkinan berkontribusi pada ekspresi gen yang berlebihan (Paulsen et al. 2018 ; Zhu et al. 2021 ). Oleh karena itu, masuk akal bahwa ekspresi berlebihan gen yang terkait dengan serat otot yang didorong oleh eccDNA berkontribusi pada peningkatan kepadatan serat otot, sehingga meningkatkan kekerasan otot.
Selain itu, otot ikan mas rumput renyah mengandung kolagen yang lebih tinggi (Ma et al. 2020 ; Tang et al. 2024 ). Khususnya, dibandingkan dengan otot ikan mas rumput biasa, eccDNA gen kolagen tipe I dan tipe II secara signifikan lebih banyak di otot ikan mas rumput renyah, sejalan dengan pengamatan ini. Menariknya, penelitian sebelumnya telah menunjukkan bahwa gen kolagen diekspresikan secara tinggi pada tingkat mRNA dan tingkat protein di otot ikan mas rumput renyah (Yu, Xie et al. 2014 ; Yu, Liu et al. 2014 ; Yu et al. 2017 ). Pada ikan mas rumput yang diberi makan kacang faba, gen yang mengkode kolagen tipe I dan tipe II tampaknya memainkan peran penting dalam pengerasan otot (Yu, Xie et al. 2014 ; Yu, Liu et al. 2014 ). Temuan ini menunjukkan bahwa peningkatan kekencangan otot rangka pada ikan mas rumput renyah kemungkinan disebabkan oleh peningkatan kadar eccDNA gen kolagen.
Kepadatan miosin berfilamen diketahui meningkat oleh kalsium (Herrera et al. 2002 ). Selain itu, peningkatan kepadatan miosin berfilamen telah dikaitkan dengan peningkatan kekencangan otot (Hatae et al. 1990 ). Khususnya, penelitian sebelumnya melaporkan kepadatan miosin berfilamen yang lebih tinggi dan kandungan kalsium yang meningkat (Liu et al. 2011 ) pada ikan mas rumput renyah dibandingkan dengan ikan mas rumput biasa, yang membantu menjelaskan mekanisme yang mendasari peningkatan kekencangan otot yang diamati pada ikan mas rumput renyah (Lin et al. 2009 ). Selain itu, banyak gen yang terkait dengan metabolisme kalsium, seperti protein kadherin (Cad), kalretikulin (CRT) dan kalmodulin (CaM), meningkat pada tingkat mRNA dan tingkat protein pada ikan mas rumput renyah (Yu, Xie et al. 2014 ; Yu, Liu et al. 2014 ; Yu et al. 2017 ). Selain itu, eccDNA dari CRT dan CaM berlimpah dalam sel otot ikan mas rumput yang sedang melangsingkan (He et al. 2024 ). Secara konsisten, eccDNA dari CRT, CaM, dan Cad juga meningkat dalam ikan mas rumput renyah, yang menunjukkan bahwa eccDNA dari gen protein yang bergantung pada kalsium sebagian berkontribusi pada peningkatan kadar kalsium, yang berkontribusi pada peningkatan kekencangan otot.
Studi terkini mengungkapkan bahwa eccDNA adalah imunostimulan bawaan yang poten, dengan kemampuan kuat untuk menginduksi produksi sitokin (Wang et al. 2021 ; Zuo et al. 2022 ). eccDNA murni atau DNA sirkular sintetis menunjukkan efek imunostimulasi yang jauh lebih kuat dibandingkan dengan DNA linier, yang secara signifikan meningkatkan ekspresi gen terkait imun (Wang et al. 2021 ). Sejalan dengan ini, analisis kami mengungkapkan bahwa mayoritas gen terkait eccDNA yang meningkat pada ikan mas rumput renyah diperkaya dalam kelas terkait imun, termasuk penolakan alograf, diabetes melitus tipe I, penyakit graft-versus-host, jalur pensinyalan JAK–STAT, dan miokarditis virus. Jalur-jalur ini terkait erat dengan penyakit yang dimediasi imun, peradangan, respons imun, stres oksidatif, apoptosis sel, dan autofagi (Seif et al. 2017 ; Abboud et al. 2020 ; Eizirik et al. 2020 ; Lasrado and Reddy 2020 ), yang menunjukkan bahwa eccDNA mungkin berperan dalam memicu respons imun pada ikan. Jika penelitian di masa mendatang mengonfirmasi hipotesis ini, eccDNA yang diidentifikasi dalam penelitian ini dapat memberikan dasar untuk mengembangkan vaksin ikan dan strategi imunoterapi.
5 Kesimpulan
Sebagai kesimpulan, dengan menggunakan strategi Circle-seq dan analisis alur kerja yang ketat, kami menghasilkan basis data eccDNA baru dan dasar untuk regulasi molekuler kekencangan otot ikan. Selama peningkatan kekencangan otot, total 10.757 eccDNA ditemukan diekspresikan secara berbeda pada ikan mas rumput renyah dibandingkan dengan ikan mas rumput biasa. EccDNA ini sebagian besar diperkaya dengan gugus fungsi yang memainkan peran penting dalam regulasi molekuler pengerasan otot. EccDNA juga dapat berfungsi sebagai elemen regulasi yang mengatur amplifikasi dan ekspresi gen. Penelitian selanjutnya harus difokuskan pada penerangan mekanisme molekuler yang dengannya eccDNA memengaruhi kekencangan otot dan menyelidiki potensinya untuk meningkatkan tekstur otot pada ikan yang dibudidayakan.

