
ABSTRAK
Meskipun kelat asam fulvik-seng (Zn-FA) dapat secara efisien mengurangi akumulasi kadmium (Cd) dalam bulir padi, sedikit yang diketahui tentang dampaknya pada kadar elemen bermanfaat lainnya (seperti seng) dalam padi, bersama dengan konsentrasi optimal dan mekanisme molekuler yang mendasarinya. Tujuan dari penelitian ini adalah untuk menyelidiki pengaruh Zn-FA terhadap akumulasi Cd dalam padi dan mekanisme metabolismenya melalui percobaan pot. Hasilnya menunjukkan bahwa perlakuan Zn-FA secara signifikan mengurangi Cd dan meningkatkan kandungan Zn dalam bulir padi, dengan 10 g/L Zn-FA diidentifikasi sebagai konsentrasi optimal untuk penyemprotan. Perlakuan Zn-FA terutama mengurangi translokasi Cd dari batang ke bulir, sehingga membatasi akumulasi Cd dalam bulir. Analisis metabolomik mengungkapkan bahwa aplikasi Zn-FA secara signifikan mengubah profil metabolisme daun padi di bawah tekanan Cd. Pada Yixiangyou586 (YXY586), Zn-FA melibatkan peningkatan regulasi flavonoid dengan sifat antioksidan, bersama dengan jalur terkait seperti biosintesis flavonoid, flavon, dan biosintesis flavonol; demikian pula, pada Gengxiangyou703 (GXY703), Zn-FA memodulasi metabolisme flavonoid. Di bawah aplikasi Zn-FA, pengurangan kandungan Cd dan peningkatan akumulasi Zn dalam bulir padi terutama terkait dengan perubahan metabolisme fenolik, terutama flavonoid dalam daun padi. Temuan penelitian ini menawarkan wawasan baru tentang mekanisme Zn-FA dalam mengurangi kandungan Cd dalam padi, sehingga memberikan landasan ilmiah dan dukungan teknis untuk memastikan produksi tanaman yang aman dan meningkatkan keamanan ekologis.
1 Pendahuluan
Kadmium (Cd) adalah logam berat beracun yang menimbulkan risiko besar bagi tanaman dan kesehatan manusia (Guo et al. 2021 ). Aktivitas antropogenik, seperti penambangan, peleburan, dan pemberian pupuk, telah berkontribusi terhadap kontaminasi Cd di tanah pertanian, sehingga menimbulkan masalah lingkungan global (Zhao et al. 2014 ). Sebaliknya, seng (Zn) merupakan mikronutrien esensial dan berperan penting dalam menjaga kesehatan manusia. Di Tiongkok dan Asia Tenggara, beras, tanaman pangan pokok, merupakan sumber utama Zn dan Cd dalam makanan manusia (Jena et al. 2023 ; Chasapis et al. 2020 ; Yang et al. 2020 ). Asupan Cd yang berlebihan dapat menyebabkan konsekuensi kesehatan yang serius, seperti kerusakan tulang, penyakit ginjal, dan peningkatan risiko kanker (Xue et al. 2023 ). Demikian pula, defisiensi Zn dalam makanan juga dapat menyebabkan berbagai masalah kesehatan (Jena et al. 2023 ; Wang et al. 2021 ), seperti pertumbuhan terhambat, kehilangan nafsu makan, gangguan fungsi imun, kerusakan DNA, dan peningkatan risiko kanker (Liu et al. 2024 ; Wessels dan Rink 2020 ). Akibatnya, mengurangi konsentrasi Cd sekaligus meningkatkan kadar Zn dalam bulir padi sangat penting untuk meningkatkan kesehatan dan gizi manusia.
Baru-baru ini, berbagai metode remediasi fisik, kimia, dan biologis telah dikembangkan untuk mengurangi kontaminasi Cd pada padi. Meskipun metode ini efektif, metode ini sering kali melibatkan kelemahan yang signifikan, seperti biaya remediasi yang tinggi dan potensi pencemaran lingkungan sekunder (Song et al. 2023 ). Sebaliknya, teknologi aplikasi daun telah menarik perhatian yang cukup besar karena efektivitas, keamanan, dan efisiensi biaya (Wang, Liu, et al. 2024 ). Sejumlah penelitian telah mengonfirmasi bahwa penyemprotan daun dengan EDTA-Zn atau ZnSO 4 secara signifikan mengurangi kandungan Cd pada padi (Zheng et al. 2023 ; Wang et al. 2020 ). Namun, aplikasi ZnSO 4 daun yang berlebihan dapat mengakibatkan efek samping, seperti daun terbakar, perkembangan malai tidak lengkap, dan hasil gabah berkurang (Xu et al. 2022 ). Lebih jauh lagi, aplikasi eksogen agen khelasi kimia seperti EDTA dapat memberikan dampak negatif pada pertumbuhan tanaman dan kualitas tanah (Wang et al. 2020 ; Han et al. 2020 ).
Zat humik, yang dikenal sebagai amandemen tanah yang hemat biaya dan ramah lingkungan, telah terbukti secara efektif mengurangi penyerapan dan akumulasi Cd dalam bulir padi (Yu et al. 2018 ). Asam fulvik (FA), suatu komponen asam humik, memiliki berat molekul yang lebih rendah dan jumlah gugus fungsi yang mengandung oksigen yang lebih tinggi (misalnya, gugus hidroksil, karboksil, fenolik, kuinon, karbonil, dan semikuinon) dibandingkan dengan asam humik. Gugus fungsi ini dapat mengikat logam berat, membentuk kompleks yang mengurangi penyerapan logam berat oleh tanaman (Gao et al. 2022 ; Wang et al. 2019 ). Misalnya, penyemprotan FA melalui daun meningkatkan pertumbuhan di bawah tekanan Cd sementara secara bersamaan mengurangi kandungan Cd dalam jaringan selada (Wang et al. 2019 ). Selain itu, penelitian kami sebelumnya menunjukkan bahwa Zn-FA adalah senyawa Zn kelat organik yang sangat efektif dalam mengurangi konsentrasi Cd dalam bulir padi (Lu et al. 2024 ). Namun, dampaknya terhadap kadar Zn dalam beras, bersama dengan konsentrasi optimal dan mekanisme molekuler yang mendasarinya, masih belum jelas.
Dalam beberapa tahun terakhir, penerapan metabolomik tanaman untuk mempelajari respons metabolik tanaman di bawah tekanan logam berat telah menarik perhatian yang semakin meningkat. Wu et al. ( 2024 ) dan lainnya menemukan bahwa paparan Cd menginduksi perubahan metabolomik pada daun padi, di mana kandungan asam amino, polisakarida, dan protein menurun dengan meningkatnya konsentrasi paparan, mengganggu 28 jalur metabolisme, dan akibatnya memengaruhi hasil dan kualitas tanaman. Oleh karena itu, penelitian ini akan memanfaatkan pendekatan metabolomik untuk menyelidiki efek Zn-FA pada perubahan metabolit daun pada padi, sehingga menjelaskan mekanisme akumulasi Cd pada padi.
Mengingat kesenjangan pengetahuan ini, dihipotesiskan bahwa perlakuan Zn-FA menghambat translokasi Cd ke gabah dengan memodulasi sintesis metabolit daun, sekaligus mendorong pembagian Zn. Tujuan utama dari penelitian ini adalah (1) untuk menyelidiki konsentrasi optimal Zn-FA untuk aplikasi daun, (2) untuk mengevaluasi dampak Zn-FA pada akumulasi Cd dan Zn dalam padi, dan (3) untuk menyelidiki pengaruhnya terhadap metabolit daun. Penelitian ini bertujuan untuk mengembangkan metode yang menjanjikan yang secara bersamaan mengurangi kadar Cd dan meningkatkan kandungan Zn dalam padi, sekaligus menjelaskan mekanisme potensial melalui pendekatan metabolomik.
2 Bahan dan Metode
2.1 Desain Percobaan
Tanah terkontaminasi yang digunakan untuk percobaan pot dikumpulkan dari sawah padi yang terkontaminasi Cd di wilayah pertambangan Tianzhu Ba di Provinsi Guizhou, Tiongkok (26°41′50″N, 108°54′10″E). pH tanah adalah 6,7 ± 0,09. Tanah dikeringkan dengan udara secara alami, diayak, dan dicampur secara menyeluruh sebelum digunakan. Lima kilogram tanah padi yang terkontaminasi Cd ditempatkan dalam pot plastik dan mengalami perlakuan genangan air selama 10 hari. Menurut penelitian kami sebelumnya (Lu et al. 2024 ), penyemprotan daun 10 g/L Zn-FA dapat secara signifikan mengurangi akumulasi Cd dalam padi. Dengan demikian, pot dibagi menjadi empat kelompok perlakuan: kelompok kontrol (CK, disemprot dengan air deionisasi) dan tiga kelompok perlakuan Zn-FA (disemprot dengan larutan Zn-FA pada konsentrasi 5, 10, dan 15 g/L), dengan tiga kali ulangan per kelompok. Dua varietas padi lokal yang dibudidayakan secara luas, Yixiangyou586 (YXY586) dan Gengxiangyou703 (GXY703), dipilih untuk pembibitan. Bibit padi yang tumbuh seragam dipilih untuk dipindahkan, dengan dua bibit ditanam per pot. Penyemprotan daun dilakukan selama tahap anakan, pembentukan bakal buah, dan pengisian gabah menggunakan kaleng semprot pneumatik. Dosis setiap perlakuan adalah 200 mL pada tahap anakan dan 300 mL pada tahap pembentukan bulir dan pengisian gabah. Kami menggunakan film plastik untuk menutupi permukaan tanah selama penyemprotan kimia untuk mengurangi kontak antara bahan kimia dan tanah. Untuk meningkatkan daya rekat larutan pada daun, ditambahkan lima hingga delapan tetes Tween 80. Penyemprotan dilakukan pada hari tanpa angin dan hujan setelah pukul 16.00 WIB agar daun padi terbasahi secara merata.
Kedalaman air dalam pot dipertahankan sekitar 2 cm selama periode pertumbuhan padi. Sekitar 0,5 g urea dan 0,5 g pupuk majemuk diberikan pada setiap pot pada tahap pembibitan, dan 1,0 g urea dan 1,2 g pupuk majemuk juga diberikan pada setiap pot pada tahap anakan, pembentukan bulir, dan pembentukan nat. Percobaan pot dilakukan di Universitas Kedokteran Guizhou (26°22′31.29″ LU, 106°38′18.38″ BT).
2.2 Pengumpulan dan Penentuan Sampel
Setelah padi matang, tiga helai daun segar dengan ujung yang mengembang penuh dikumpulkan, dibersihkan dengan air deionisasi, ditempatkan dalam pendingin es kering, dan segera dipindahkan ke lemari es pada suhu -80°C untuk pengawetan guna analisis metabolomik. Gabah, akar, batang, dan daun dari seluruh tanaman padi dikumpulkan secara terpisah, dibersihkan dengan air deionisasi, dan dibawa kembali ke laboratorium untuk diproses. Sampel-sampel tersebut pertama-tama mengalami pendinginan selama 30 menit pada suhu 105°C, diikuti dengan pengeringan pada suhu 60°C hingga mencapai berat konstan. Sekam dan beras merah dipisahkan menggunakan mesin pengupas kecil. Sampel akar, batang, daun, dan gabah kemudian digiling menjadi bubuk, kemudian disaring melalui saringan 100 mesh, dan disimpan untuk analisis. Sampel tanah dikeringkan dengan udara, digiling menggunakan mortar keramik, diproses melalui saringan 100 mesh, dan disimpan untuk penggunaan selanjutnya.
Prosedur pencernaan sampel mengikuti protokol yang ditetapkan oleh Lu et al. Untuk sampel tanaman, 0,1 g sampel dicerna dengan 3 mL HNO3 . Selanjutnya, sampel dicerna pada suhu 158°C selama 16 jam. Satu mililiter 30% H2O2 ditambahkan ke dalam campuran dan kemudian dipanaskan pada pelat pemanas pada suhu 95° C selama 1 jam. Suhu kemudian dinaikkan menjadi 105°C untuk menguapkan asam. Untuk sampel tanah, 0,05 g sampel ditimbang ke dalam wadah Teflon, dicerna dengan 3 mL HNO3 dan 1 mL HF, dan kemudian dicerna pada suhu 158°C selama 48 jam. Satu mililiter HNO3 ditambahkan ke dalam campuran dan kemudian diuapkan pada pelat panas pada suhu 105°C. Semua larutan dipindahkan dan diencerkan untuk analisis. Konsentrasi Cd dan Zn diukur dengan spektrometer massa plasma induktif (NexION 2000, PerkinElmer Co. Ltd., AS).
Selama analisis, blanko metode, bahan referensi bersertifikat, dan sampel duplikat digunakan untuk jaminan kualitas dan pengendalian kualitas. Simpangan baku relatif (RSD) duplikat kurang dari 5%. Bahan bersertifikat berupa daun jeruk (GBW10020) dan tanah kuning-merah (GSS-5a) digunakan untuk sampel tanaman dan tanah, dengan perolehan kembali berkisar antara 106,9% hingga 116,9% dan 87,2% hingga 98,3%, masing-masing.
2.3 Perhitungan Faktor Transfer
Dalam penelitian ini, faktor transfer (TF) Cd dan Zn dari akar ke gabah, batang ke gabah, daun ke gabah, dan batang ke daun dihitung menurut metode yang diuraikan oleh Zheng et al. ( 2023 ). Rumus perhitungan khususnya adalah sebagai berikut:

Dalam rumus ini, C butir , C akar , C batang , dan C daun masing-masing mewakili konsentrasi Cd atau Zn dalam butir, akar, batang, dan daun.
2.4 Analisis Statistik
Dalam penelitian ini, data diolah dengan menggunakan Microsoft Excel 2019. Semua data dinyatakan dalam bentuk mean ± simpangan baku (
± s, n = 3), analisis varians satu arah (ANOVA) dan grafik dilakukan menggunakan perangkat lunak GraphPad Prism 9.5.0 dengan tingkat signifikansi 0,05.
2.5 Analisis Metabolomik Tak Tertarget
2.5.1 Ekstraksi Sampel
Sampel daun padi sebanyak 100 mg ditambahkan ke tabung sentrifus 2 mL dan manik penggiling berdiameter 6 mm ditambahkan. Selanjutnya, 800 μL larutan ekstraktan (metanol: air = 4:1 v:v) ditambahkan, diikuti dengan ekstraksi ultrasonik pada suhu rendah selama 30 menit. Sampel kemudian ditempatkan pada suhu -20°C selama 30 menit, setelah itu disentrifus selama 15 menit (4°C, 13.000 g). Supernatan dipindahkan ke vial injeksi untuk analisis LC–MS/MS, yang dilakukan pada sistem Thermo UHPLC-Q Exactive di Majorbio Bio-Pharm Technology Co. Ltd. (Shanghai, Tiongkok). Pemisahan kromatografi dilakukan menggunakan kolom ACQUITY UPLC BEH C18 (100 mm × 2,1 mm id, 1,7 μm; Waters, AS) dengan fase mobil yang terdiri dari pelarut A yang mengandung 2% asetonitril dalam air (mengandung 0,1% asam format) dan pelarut B yang terdiri dari asetonitril (mengandung 0,1% asam format). Volume injeksi adalah 3 μL, suhu kolom diatur pada 40°C, dan laju alir diatur pada 0,4 mL/menit. Prosedur elusi gradien adalah seperti yang diuraikan di bawah ini: 0–0,5 menit, 2% pelarut B; 0,5–7,5 menit, 2%–35% pelarut B; 7,5–13 menit, 35%–95% pelarut B; 13–14,4 menit, 95% pelarut B; 14,4–14,5 menit, 95%–2% pelarut B; 14,5–16 menit, 2% pelarut B. Mode ion positif dan negatif digunakan selama analisis spektrometri massa, dengan rentang pindaian dari 70 hingga 1050 m/z dan voltase masing-masing 3500 V dan 3000 V. Selama analisis instrumental, sampel kontrol kualitas (QC), yang disiapkan dengan mencampur semua sampel dengan volume yang sama, dimasukkan setelah setiap 12 sampel analitis untuk memantau stabilitas analisis.
2.5.2 Analisis Data Metabolomik
Data mentah LC/MS disaring dari garis dasar, diintegrasikan, puncak diidentifikasi, diselaraskan, dan waktu retensi dikoreksi menggunakan perangkat lunak Progenesis QI (Waters, Milford, AS), dan informasi spektral massa MS dan MS/MS kemudian dicocokkan dengan basis data metabolit spesifik yang dibangun sendiri (MJDBPM, Shanghai Majorbio, Tiongkok) untuk memperoleh informasi metabolit. Semua fitur dengan simpangan baku lebih besar dari 30% berdasarkan sampel QC dikecualikan untuk menghilangkan interferensi. Data kemudian dievaluasi menggunakan analisis komponen utama (PCA) dan analisis diskriminan kuadrat terkecil parsial (PLS-DA) oleh paket R “ropls (versi 1.6.2).” Merujuk pada metode penelitian sebelumnya, menggunakan uji Student, metabolit dengan Variabel penting dalam proyeksi (VIP) > 1 dan nilai- p < 0,05 dianggap sebagai metabolit yang berbeda secara signifikan (Wang, Mu, et al. 2024 ; Wang, Mu, et al. 2023 ; Xiong et al. 2023 ). Anotasi metabolit diferensial dari jalur metabolisme dilakukan melalui basis data Kyoto Encyclopedia of Genes and Genomes (KEGG). Metabolit ini diklasifikasikan menurut fungsi biologis dan keterlibatannya dalam jalur metabolisme. Analisis pengayaan jalur dilakukan menggunakan paket Python “scipy stats” dan jalur biologis yang paling relevan dengan perlakuan eksperimental diidentifikasi melalui uji eksak Fisher.
3 Hasil
3.1 Pengaruh Penyemprotan Zn-FA terhadap Akumulasi Cd pada Jaringan Padi
Efek aplikasi daun pada Zn-FA mengurangi kandungan Cd dalam bulir padi untuk kedua varietas padi, seperti yang ditunjukkan pada Gambar 1. Untuk YXY586 (Gambar 1a ), kandungan Cd dalam bulir padi menurun sebesar 34,1%, 64,8%, dan 56,0% pada perlakuan dengan 5, 10, dan 15 g/L Zn-FA, masing-masing, dengan semua sampel yang diberi perlakuan menunjukkan kadar Cd di bawah batas yang ditetapkan Cina untuk padi (0,2 mg/kg). Penurunan kandungan Cd terbesar diamati pada konsentrasi 10 g/L relatif terhadap kelompok kontrol. Selain itu, aplikasi Zn-FA memengaruhi penyerapan Cd pada akar, batang, dan daun padi. Pada perlakuan 5, 10, dan 15 g/L Zn-FA, kandungan Cd menurun sebesar 24,3%, 23,1%, dan 10,8% pada akar; 16,3%, 14,3%, dan 44,9% pada batang; dan 21,7%, 21,7%, dan 34,8% pada daun, berturut-turut. Untuk GXY703 (Gambar 1a ), kandungan Cd pada gabah padi menurun sebesar 25,6%, 32,1%, dan 17,9% pada perlakuan dengan 5, 10, dan 15 g/L Zn-FA, berturut-turut, relatif terhadap kelompok kontrol. Penurunan kandungan Cd yang paling menonjol pada gabah dicapai pada konsentrasi 10 g/L. Pada varietas ini, kandungan Cd pada akar dan batang lebih rendah dibandingkan pada kelompok kontrol pada 5 g/L Zn-FA tetapi meningkat pada konsentrasi yang lebih tinggi. Kandungan Cd dalam daun menunjukkan penurunan yang signifikan pada ketiga konsentrasi perlakuan ( p < 0,0001), yaitu masing-masing menurun sebesar 59,4%, 53,6%, dan 68,8% dibandingkan dengan kelompok kontrol.
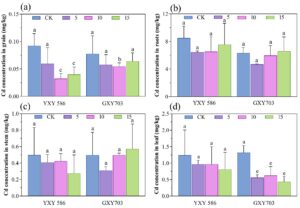
3.2 Pengaruh Penyemprotan Zn-FA terhadap Akumulasi Zn pada Jaringan Padi
Pengaruh pemberian Zn-FA melalui daun terhadap kandungan Zn dalam jaringan padi ditunjukkan pada Gambar 2. Pada varietas YXY586, dibandingkan dengan kelompok kontrol, kandungan Zn dalam gabah padi meningkat sebesar 18,3% dan 25,7% pada perlakuan Zn-FA 10 dan 15 g/L, sedangkan kandungan Zn dalam gabah padi menurun sebesar 6,1% pada perlakuan Zn-FA 5 g/L. Selain itu, pada tiga perlakuan konsentrasi, kandungan Zn dalam batang meningkat sebesar 12,9%, 65,1%, dan 14,2%, diikuti oleh peningkatan sebesar 3,2%, 33,6%, dan 28,0% pada daun. Pada varietas GXY 703, relatif terhadap kelompok kontrol, kandungan Zn dalam bulir padi meningkat sebesar 15,6%, 23,7%, dan 24,4% pada perlakuan Zn-FA 5, 10, dan 15 g/L. Kandungan Zn dalam akar dan batang padi juga meningkat pada perlakuan 10 dan 15 g/L, dengan kandungan Zn meningkat sebesar 7,4% dan 11,6% pada akar, diikuti oleh peningkatan yang lebih tinggi sebesar 16,3% pada batang pada kedua perlakuan.
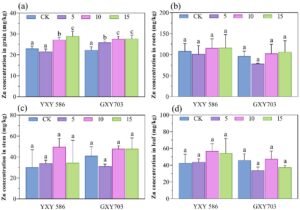
3.3 Dampak Aplikasi Zn-FA pada Daun terhadap Faktor Translokasi Logam
Nilai faktor transfer (TF) yang dihitung disajikan dalam Gambar 3. Ini dengan jelas menunjukkan bahwa perlakuan Zn-FA memengaruhi translokasi Cd pada kedua varietas padi, tetapi dampak pada translokasi Zn dalam organ kedua varietas padi tidak signifikan (Gambar S1 ). Pada varietas YXY586, dibandingkan dengan kelompok kontrol, pengurangan utama dalam translokasi Cd diamati dari akar, batang, dan daun ke gabah padi, yang mengakibatkan penurunan kandungan Cd dalam gabah padi. Pada varietas GXY703, pengurangan utama dalam translokasi Cd terjadi dari batang ke daun. Selain itu, pada konsentrasi 10 dan 15 g/L, Zn-FA secara signifikan menghambat translokasi Cd dari akar dan batang ke gabah padi.

3.4 Analisis Metabolomik
Berdasarkan partial least squares discriminant analysis (PLS-DA), pemisahan yang jelas diamati antara aplikasi daun dengan konsentrasi reagen yang berbeda dan kelompok kontrol untuk kedua varietas (YXY586 dan GXY703) (Gambar 4 ). Temuan ini menunjukkan bahwa aplikasi Zn-FA berdampak pada metabolit daun padi. Dengan menggunakan kriteria skor VIP (VIP > 1) dari model OPLS-DA serta uji-t Student ( p < 0,05), total 984 metabolit diferensial diidentifikasi.

Seperti yang ditunjukkan pada Gambar 5 , pada varietas YXY586, dibandingkan dengan kontrol, 491 metabolit diferensial berubah secara signifikan setelah penyemprotan 5 g/L Zn-FA (238 mengalami peningkatan dan 253 mengalami penurunan). Sebanyak 339 metabolit berubah secara signifikan setelah penyemprotan 10 g/L Zn-FA (189 mengalami peningkatan dan 150 mengalami penurunan), dan 129 metabolit berubah secara signifikan setelah penyemprotan 15 g/L Zn-FA (55 mengalami peningkatan dan 74 mengalami penurunan). Demikian pula, pada varietas GXY703, 153 metabolit berubah secara signifikan setelah penyemprotan 5 g/L Zn-FA (53 mengalami peningkatan dan 100 mengalami penurunan), diikuti oleh 122 metabolit berubah secara signifikan setelah penyemprotan 10 g/L Zn-FA (49 mengalami peningkatan dan 73 mengalami penurunan), dan 140 metabolit (60 mengalami peningkatan dan 80 mengalami penurunan) berubah secara signifikan setelah penyemprotan dengan 15 g/L Zn-FA.
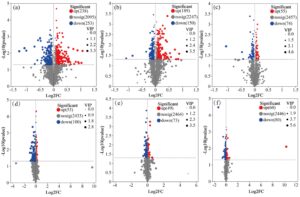
Pada varietas YXY586, setelah perlakuan 5 dan 10 g/L, metabolit yang meningkat terutama fenolik (termasuk flavonoid) dan turunannya, sedangkan metabolit yang menurun terutama meliputi terpena, lipid, asam organik, dan turunannya. Setelah perlakuan 15 g/L, lipid dan terpenoid terutama meningkat, sedangkan fenolik (termasuk flavonoid) dan turunannya, asam organik, dan turunannya terutama menurun. Demikian pula, pada varietas GXY703, setelah perlakuan 5 dan 10 g/L, metabolit yang meningkat terutama fenolik (termasuk flavonoid) dan turunannya, sedangkan metabolit yang menurun terutama meliputi terpenoid, asam organik, dan turunannya, dan lipid. Setelah perlakuan 15 g/L, terpena dan lipid terutama meningkat, sedangkan flavonoid, karbohidrat, dan turunannya terutama menurun.
Hasil analisis pengayaan jalur KEGG dari metabolit diferensial ditunjukkan pada Gambar 6. Pada varietas YXY586, 40 jalur metabolisme diidentifikasi dalam kelompok kontrol versus 5 g/L Zn-FA, dengan efek signifikan ( p < 0,05) yang diamati dalam metabolisme asam α-linolenat, biosintesis fenilpropanoid, biosintesis flavonoid, biosintesis flavon dan flavonol, dan metabolisme triptofan. Jumlah jalur metabolisme terbesar diidentifikasi dalam kelompok kontrol versus 10 g/L Zn-FA, dengan total 42 jalur metabolisme yang diidentifikasi. Di antara jalur-jalur ini, biosintesis kofaktor, biosintesis flavonoid, biosintesis flavon dan flavonol, metabolisme sisteina dan metionina, dan metabolisme asam arakidonat terpengaruh secara signifikan ( p < 0,05). Pada kelompok kontrol versus 15 g/L Zn-FA, total 24 jalur metabolisme diidentifikasi, termasuk biosintesis fenilpropanoid, fenilalanina, tirosin, dan biosintesis triptofan, metabolisme askorbat dan aldarat, biosintesis berbagai alkaloid, dan metabolisme tirosin, yang secara signifikan dipengaruhi ( p < 0,05). Temuan ini menunjukkan bahwa variasi dalam konsentrasi Zn-FA secara substansial mempengaruhi jalur metabolisme pada daun padi. Perlakuan Zn-FA 10 g/L mempengaruhi jalur metabolisme paling banyak pada daun padi, yang menunjukkan bahwa konsentrasi ini memberikan dampak terbesar pada metabolisme daun. Selain itu, biosintesis flavon dan flavonol, serta biosintesis flavonoid, umumnya dipengaruhi oleh perlakuan Zn-FA 5 dan 10 g/L.

Pada varietas GXY703, 28 jalur metabolisme diidentifikasi dalam kelompok kontrol versus 5 g/L Zn-FA, dengan efek signifikan ( p < 0,05) yang diamati dalam metabolisme tirosin, metabolisme asam amino siano, biosintesis flavon dan flavonol, biosintesis aminoasil-tRNA, metabolisme gliserofosfolipid, metabolisme askorbat dan aldarat, metabolisme asam amino D, biosintesis berbagai alkaloid, biosintesis glukosinolat, dan transduksi sinyal hormon tanaman. Demikian pula, total 25 jalur diidentifikasi dalam kelompok kontrol versus 10 g/L Zn-FA, dengan efek signifikan pada metabolisme asam amino D, metabolisme tirosin, dan transduksi sinyal hormon tanaman ( p < 0,05). Sedangkan untuk kelompok kontrol versus 15 g/L Zn-FA, terdeteksi 27 jalur, dengan efek signifikan ( p < 0,05) pada metabolisme fenilalanina, biosintesis fenilpropanoid, biosintesis kutin, suberina dan lilin, biosintesis fenilalanina, tirosin, dan triptofan, biosintesis aminoasil-tRNA, metabolisme askorbat dan aldarat, metabolisme asam amino D, biosintesis ubikuinon dan terpenoid-kuinon lainnya, dan biosintesis alkaloid tropan, piperidin, dan piridin. Hasil ini menunjukkan bahwa berbagai konsentrasi Zn-FA memiliki efek berbeda pada jalur metabolisme pada daun padi. Selain itu, metabolisme tirosin dan metabolisme asam amino D merupakan jalur metabolisme umum yang dipengaruhi oleh perlakuan Zn-FA 5 dan 10 g/L.
4 Diskusi
Hasilnya menunjukkan bahwa Zn-FA efektif mengurangi konsentrasi Cd dalam bulir padi dari kedua varietas pada tingkat yang bervariasi, dengan pengurangan paling signifikan dalam konsentrasi Cd diamati pada konsentrasi 10 g/L. Namun, dampak pada kandungan Cd dalam jaringan lain bervariasi. Penelitian sebelumnya menunjukkan bahwa perbedaan genetik di antara varietas padi, serta konsentrasi Zn yang diberikan, dapat memengaruhi distribusi Cd di berbagai jaringan (Zhen et al. 2021 ; Zhao et al. 2021 ). Penelitian telah menunjukkan bahwa aplikasi pupuk Zn melalui daun mengurangi kandungan logam berat pada bagian tanaman yang dapat dimakan seperti padi dan gandum (Zheng et al. 2023 ; Saifullah et al. 2016 ; Yang et al. 2011 ).
Efek ini disebabkan oleh sifat kimia, jalur serapan, dan mekanisme transpor Zn dan Cd yang serupa. Sebagai kation divalen, Zn berbagi sistem transpornya dengan Cd. Ketika keduanya hadir, mereka bersaing untuk tempat pengikatan pada transporter; konsentrasi Zn yang tinggi dapat mengikat lebih erat pada transporter, sehingga mengurangi efisiensi transfer Cd ke endosperma sambil meningkatkan transfer Zn (Zheng et al. 2024 ), seperti melalui transporter OsIRT1 , OsHMA2 , OsHMA3 , dan OsZIP7 . Peningkatan ekspresi OsIRT1 meningkatkan penyerapan Zn pada padi, sementara penurunan ekspresi OsHMA2 menurunkan konsentrasi Cd dalam xilem. OsHMA3 mengatur pengangkutan Cd xilem padi dengan memediasi segregasi vesikular Cd di sel akar, sedangkan OsZIP7 memfasilitasi pemuatan xilem di akar dan transfer antar pembuluh di nodus, terutama dalam menyalurkan Zn dan Cd ke jaringan yang sedang berkembang dan bulir padi (Zheng et al. 2023 ; Hu et al. 2019 ).
Lebih jauh lagi, Zn pada daun dapat mengurangi konsentrasi glutathione (GSH) pada daun dan batang bendera, mengurangi ekspresi gen transporter Zn, dan membatasi transpor Cd pada xilem, sehingga mendorong fiksasi Cd pada bagian tanaman lain dan akhirnya mengurangi kandungan Cd pada gabah padi (Zheng et al. 2023 ). Selain itu, pemberian Zn pada daun menghambat peningkatan hidrogen peroksida (H 2 O 2 ) dan malondialdehid (MDA) yang diinduksi Cd, meningkatkan aktivitas superoksida dismutase (SOD) dan katalase (CAT), dan mengurangi kerusakan oksidatif. Peningkatan ini meningkatkan pertumbuhan dan karakteristik fotosintesis tanaman, sehingga mengurangi kerusakan yang disebabkan oleh stres Cd (Faizan et al. 2021 ). Mirip dengan penelitian ini, penyemprotan daun 0,5% Zn-EDTA dapat secara signifikan mengurangi kandungan Cd dalam biji-bijian, mungkin karena Zn-EDTA menghambat penyerapan Cd oleh sistem akar padi dan meningkatkan akumulasi Zn dalam biji-bijian padi, sehingga mengurangi translokasi Cd ke biji-bijian (Wang et al. 2020 ). Selain itu, Wang, Li, et al. ( 2023 ) melaporkan bahwa penerapan FA dapat secara signifikan mengurangi dampak buruk stres Cd pada perkecambahan biji jagung dan pertumbuhan bibit. Pengurangan kandungan Cd oleh Zn-FA mungkin karena banyaknya gugus fungsi, seperti gugus alifatik, hidroksil, amida, kuinon, keton, dan karbonil, yang dapat bereaksi dengan ion logam untuk membentuk kompleks yang stabil, sehingga mengurangi kandungan Cd dalam tanaman (Song et al. 2023 ). Akibatnya, aplikasi Zn-FA melalui daun tampaknya menjadi strategi yang menjanjikan untuk mengurangi akumulasi Cd pada bulir padi, kemungkinan karena efek sinergis dari FA dan Zn. Penelitian saat ini dilakukan hanya pada tanah di Provinsi Guizhou, dan perbedaan sifat fisikokimia dari tanah yang berbeda dapat secara signifikan mempengaruhi efektivitas aplikasi Zn-FA; oleh karena itu, perlu untuk menindaklanjuti dengan uji pot untuk membandingkan beberapa jenis tanah. Untuk kedua varietas tersebut, aplikasi Zn-FA menghasilkan peningkatan kandungan Zn pada bulir padi. Efek ini konsisten dengan penelitian sebelumnya yang menunjukkan bahwa aplikasi Zn melalui daun secara efektif meningkatkan kandungan Zn pada tanaman. Misalnya, Duan et al. ( 2018 ) melaporkan peningkatan kandungan Zn sebesar 90,5% pada padi setelah aplikasi ZnSO4 melalui daun . Selain itu, temuan kami menunjukkan korelasi positif antara konsentrasi Zn yang diaplikasikan melalui daun dan kandungan Zn pada bulir padi (Gambar 2a ). Tren ini sejalan dengan hasil yang dilaporkan oleh Zheng et al. ( 2023)), di mana kandungan Zn meningkat sebesar 93% dan 172% dalam gabah padi di bawah perlakuan dengan Zn 0,2% dan 0,4%, berturut-turut. Penelitian telah menunjukkan bahwa Zn yang diaplikasikan melalui daun diserap secara efisien oleh daun dan diangkut ke gabah, sehingga meningkatkan akumulasi Zn dalam gabah (Ahmad et al. 2025 ). Setelah Cd diserap oleh sistem akar, ia ditranslokasi ke batang melalui xilem, difasilitasi oleh transporter seperti OsHMA2 dan OsHMA3. Cd terutama terakumulasi dalam gabah melalui floem, ditranslokasi dari batang ke gabah (Wiggenhauser et al. 2021 ; Zhong et al. 2021 ). Varietas padi yang berbeda menunjukkan kapasitas yang berbeda-beda untuk akumulasi dan transportasi Cd di akar, batang, daun, dan malai mereka. Batang memainkan peran penting dalam mengangkut unsur-unsur mineral ke berbagai organ, dengan translokasi Cd ke atas terutama dibatasi pada ruas batang (Lu et al. 2023 ). Dalam penelitian ini, aplikasi Zn-FA melalui daun mengurangi translokasi Cd dari batang ke gabah pada kedua varietas padi, kecuali varietas GXY703 yang diobati dengan 5 g/L Zn-FA, yang akibatnya mengurangi akumulasi Cd pada gabah. Zheng et al. ( 2023 ) melaporkan bahwa aplikasi Zn melalui daun meningkatkan sekuestrasi Cd di dalam batang padi dengan menurunkan ekspresi gen transporter Zn di dalam daun bendera, sehingga mengurangi translokasi Cd melalui xilem.
Studi ini menunjukkan bahwa aplikasi Zn-FA pada daun mengubah profil metabolik daun padi, khususnya dengan mengatur senyawa fenolik, termasuk flavonoid, yang merupakan subkelas fenolik. Fenolik adalah metabolit sekunder penting yang dibutuhkan untuk sintesis lignin dan pigmen tanaman (Elguera et al. 2012 ). Tanaman mengurangi efek toksik logam pada tingkat sel dengan mengeluarkan fenolik, yang mencegah pecahnya membran dan kerusakan oksidatif, proses yang pada akhirnya dapat memengaruhi transfer elektron dan bahkan menyebabkan kematian sel, berfungsi sebagai mekanisme perlindungan diri (Okem et al. 2015 ). Sementara itu, laporan telah menunjukkan bahwa senyawa fenolik juga dapat mengurangi toksisitas dalam sitoplasma dengan mengkelat atau mengkomplekskan dengan logam untuk membentuk logam yang diendapkan. Dengan demikian, sifat antioksidan dan khelasi logam dari senyawa fenolik memainkan peran penting dalam melindungi tanaman dari stres yang disebabkan oleh logam (Jańczak-Pieniążek et al. 2022 ).
Dalam penelitian ini, fenolik seperti 6-caffeoylsucrose, isoferuloyl C1-glucuronide, dan asam dattelic masing-masing meningkat sebesar 1,14, 1,39, dan 1,29 kali lipat, pada perlakuan Zn-FA 5 g/L dalam varietas YXY586. Demikian pula, asam 3-feruloylquinic, 4′-O-galloyl sukrosa, dan 6-caffeoyl sukrosa meningkat sebesar 1,11, 1,14, dan 1,14 kali lipat pada perlakuan Zn-FA 10 g/L. Akan tetapi, pada Zn-FA 15 g/L, senyawa fenolik menurun, yang menunjukkan bahwa konsentrasi Zn-FA yang tinggi dapat menekan biosintesis fenolik. Pada varietas GXY703, 1-O-Cinnamoyl-beta-D-gentiobiose meningkat 1,07 kali lipat pada perlakuan Zn-FA 5 g/L, sementara asam dihidroferulat, asam 4-kumarat, dan asam kumarat meningkat 1,04, 1,12, dan 1,22 kali lipat pada perlakuan Zn-FA 10 g/L. Ester asam tartarat monoferuloil menurun 0,95 kali lipat pada perlakuan Zn-FA 15 g/L, kemungkinan karena penghambatan sintesisnya yang disebabkan oleh konsentrasi tinggi Zn-FA 15 g/L.
Di antara senyawa fenolik, flavonoid, subkelas fenolik, secara signifikan dipengaruhi oleh perlakuan Zn-FA. Flavonoid adalah senyawa polifenol yang terjadi secara alami, berfungsi dalam mengatur pertumbuhan sel dan memberikan resistensi terhadap stres biotik dan abiotik. Di bawah stres abiotik, sintesis dan akumulasi flavonoid meningkat (Li et al. 2022 ). Selain itu, penelitian telah menunjukkan bahwa flavonoid melindungi sel dari kerusakan oksidatif melalui pembersihan radikal bebas, peningkatan aktivitas antioksidan non-enzimatik, dan pengaturan keseimbangan redoks. Selain itu, flavonoid menunjukkan banyak aktivitas biologis, seperti antitumor, antioksidan, dan antiinflamasi (Chen et al. 2023 ; Tang et al. 2023 ; Lu et al. 2018 ).
Pada varietas YXY586, dibandingkan dengan kontrol, di bawah perlakuan Zn-FA 5 dan 10 g/L, flavonoid seperti Luteolin-6-C-Glucoside meningkat sebesar 1,44 dan 1,41 kali lipat, Quercetin 3-beta-laminaribioside meningkat sebesar 1,34 dan 1,28 kali lipat, dan eriodictyol meningkat sebesar 2,10 dan 1,83 kali lipat, masing-masing. Flavonoid diketahui dapat mengkelat logam berat, dan ini dapat meningkatkan mekanisme pertahanan tanaman terhadap stres logam berat (Keilig dan Ludwig-Mueller 2009 ). Namun, pada konsentrasi yang lebih tinggi dari 15 g/L Zn-FA, penurunan kadar flavonoid diamati, yang menunjukkan bahwa Zn-FA yang berlebihan dapat menghambat sintesis flavonoid. Respons yang bergantung pada konsentrasi ini menyoroti pentingnya mengoptimalkan tingkat aplikasi Zn-FA untuk memaksimalkan efek menguntungkannya. Lebih jauh lagi, pada varietas GXY703, 3′-metoksiapigenin, turunan apigenin, meningkat 1,11 kali lipat pada perlakuan Zn-FA 5 g/L. Apigenin telah terbukti mengatur respons seluler terhadap stres oksidatif dan kerusakan DNA (Sung et al. 2016 ), yang selanjutnya mendukung peran flavonoid dalam mitigasi stres Cd.
Singkatnya, aplikasi Zn-FA pada konsentrasi optimal meningkatkan biosintesis senyawa fenolik, khususnya flavonoid, pada daun padi. Flavonoid, sebagai subkelas utama fenolik, dapat memainkan peran penting dalam mengurangi toksisitas Cd, mengurangi translokasinya, dan melindungi struktur seluler dari kerusakan oksidatif. Temuan ini tidak hanya memberikan strategi potensial untuk produksi padi yang lebih aman di lingkungan yang terkontaminasi Cd tetapi juga mengungkap kemungkinan mekanisme yang terlibat, yaitu melalui perubahan metabolisme fenolik, khususnya metabolisme flavonoid.
5 Kesimpulan
Studi ini menemukan bahwa aplikasi Zn-FA melalui daun tidak hanya menurunkan kandungan Cd dalam gabah tetapi juga meningkatkan kandungan Zn, sehingga meningkatkan kualitas gizi gabah. Konsentrasi 10 g/L Zn-FA diidentifikasi sebagai konsentrasi optimal untuk penyemprotan, dengan penurunan paling signifikan yang diamati pada konsentrasi ini, yang secara efektif menghambat translokasi Cd dari batang ke gabah. Lebih jauh lagi, aplikasi Zn-FA (5 dan 10 g/L) memodulasi profil metabolisme daun padi, khususnya dengan meningkatkan senyawa fenolik, khususnya flavonoid, yang memainkan peran penting dalam mengurangi toksisitas logam berat. Namun, aplikasi Zn-FA yang berlebihan (15 g/L) menunjukkan efek yang berlawanan dibandingkan dengan perlakuan 5 dan 10 g/L. Hal ini dapat menekan biosintesis senyawa bermanfaat ini. Secara keseluruhan, penelitian ini tidak hanya memberikan solusi potensial untuk produksi padi yang lebih aman di lingkungan yang terkontaminasi Cd, tetapi juga mengungkap bahwa Zn-FA mengurangi akumulasi Cd dalam bulir padi dengan memodulasi metabolit daun melalui perubahan metabolisme fenolik, khususnya metabolisme flavonoid. Penelitian di masa mendatang harus difokuskan pada pengoptimalan protokol aplikasi Zn-FA dan mengeksplorasi kemanjurannya di berbagai kondisi lingkungan untuk memastikan praktik pertanian yang berkelanjutan dan aman.

