
ABSTRAK
Tinjauan ini menyelidiki peran polifenol, senyawa alami yang banyak ditemukan dalam makanan, untuk memengaruhi jalur metabolisme yang terlibat dalam termogenesis dan pencoklatan jaringan adiposa putih (WAT). Banyak protein menunjukkan pola ekspresi yang berubah setelah konsumsi polifenol yang berkepanjangan, dengan peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) dikenal sebagai pengatur utama, yang berkontribusi terhadap peningkatan termogenisitas jaringan adiposa. Polifenol dapat meningkatkan aktivitas PGC-1α, merangsang pencoklatan WAT, dan meningkatkan termogenesis jaringan adiposa coklat (BAT). Berbagai kelas polifenol dieksplorasi, bersama dengan pensinyalan protein yang luas dan implikasi fisiologis dari temuan ini. Pemahaman yang komprehensif tentang berbagai protein dan jalur yang terlibat dalam studi pencoklatan dapat memberi pembaca perspektif yang lebih luas tentang respons termodulasi jaringan adiposa terhadap polifenol dan membimbing mereka ke strategi terapi inovatif untuk metabolisme lipid, obesitas, dan gangguan metabolisme terkait.
1 Pendahuluan
Jaringan adiposa, yang dulunya dianggap sebagai depot penyimpanan energi belaka, telah muncul sebagai organ endokrin yang sangat dinamis yang memainkan peran penting dalam mengatur metabolisme seluruh tubuh [ 1 ]. Di antara berbagai fungsinya, kapasitas jaringan adiposa putih (WAT) untuk mengalami “pencoklatan”—suatu proses yang ditandai dengan perolehan fitur adiposit berwarna coklat—telah menarik perhatian yang signifikan [ 2 ]. Peralihan fenotipik ini disertai dengan peningkatan pengeluaran energi dan peningkatan sensitivitas insulin, yang berkorelasi dengan status metabolisme yang lebih baik secara keseluruhan.
Jaringan adiposa coklat (BAT) didistribusikan secara strategis dalam tubuh, terutama di daerah supraklavikula, aksila, serviks, paraspinal, mediastinal, dan perut pada manusia, dan daerah intraskapular pada tikus. Meskipun melimpah pada bayi baru lahir dan bayi, massa BAT menurun seiring bertambahnya usia [ 3 ]. UCP1 terletak di membran dalam mitokondria dan merangsang termogenesis dengan menghilangkan gaya gerak proton yang dipertahankan oleh rantai transpor elektron (bertanggung jawab untuk memompa elektron keluar), jika tidak, proton akan bocor di membran dalam. Sebagai respons, untuk menjaga homeostasis, peningkatan laju masuknya substrat didorong ke mitokondria yang karenanya mengonsumsi lebih banyak energi [ 4 ]. Meskipun memiliki kesamaan fungsional, jaringan adiposa coklat dan krem muncul dari garis keturunan embrionik yang berbeda. Jaringan adiposa berwarna krem, mempunyai asal usul perkembangan yang lebih dekat dengan lemak putih, sedangkan adiposit coklat berasal dari sel progenitor yang sama dengan sel otot (miosit) [ 5 ], dan diatur oleh kromosom dan mekanisme yang berbeda [ 6 , 7 ].
Mengingat bahwa obesitas muncul akibat ketidakseimbangan kronis antara asupan dan pengeluaran energi, intervensi yang meningkatkan pengeluaran energi dianggap sebagai strategi terapi yang menjanjikan. Penelitian terbaru menunjukkan bahwa peningkatan aktivitas termogenik dalam jaringan adiposa menghadirkan jalur terapi potensial untuk mengobati obesitas pada model manusia dan tikus [ 8 – 11 ].
Polifenol, kelompok beragam senyawa yang terjadi secara alami yang ditemukan dalam makanan berbasis tanaman, telah dipelajari secara ekstensif untuk manfaat kesehatan potensialnya, termasuk sifat antioksidan, anti-inflamasi, dan anti-karsinogeniknya [ 12 – 15 ]. Penelitian terbaru telah menyoroti kemungkinan menarik bahwa polifenol juga dapat memodulasi fungsi jaringan adiposa, khususnya dengan mendorong pencoklatan. Koaktivator gamma 1-alfa reseptor proliferator peroksisom (PGC-1α) adalah pengatur utama biogenesis dan fungsi mitokondria. Ini memainkan peran sentral dalam proses pencoklatan dengan mengoordinasikan ekspresi gen dalam termogenesis. Beberapa penelitian telah menunjukkan bahwa polifenol dapat mengaktifkan PGC-1α, merangsang pencoklatan WAT dan meningkatkan termogenesis non-menggigil jaringan adiposa coklat (BAT) [ 16 , 17 ].
Tinjauan ini akan menyelidiki mekanisme molekuler yang mendasari efek polifenol pada pencoklatan jaringan adiposa dengan fokus khusus pada peran PGC-1α. Kami akan membahas berbagai kelas polifenol yang telah terbukti mendorong pencoklatan, jalur pensinyalan potensial yang terlibat, dan implikasi fisiologis dari temuan ini. Memahami mekanisme yang digunakan polifenol untuk memodulasi pencoklatan jaringan adiposa dapat memberikan strategi terapi baru untuk mengobati obesitas dan gangguan metabolik terkait.
2 Analisis VosViewer
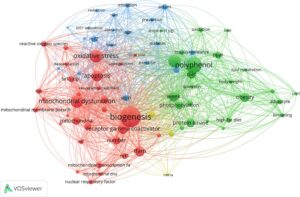
Bahasa Indonesia: Dengan menelusuri basis data PubMed untuk istilah PGC-1α (atau PGC-1alpha) dan polifenol, kami mengidentifikasi 236 artikel relevan yang diterbitkan antara tahun 2006 dan 2024. Untuk memvisualisasikan interkoneksi di antara kata-kata yang ditemukan dalam judul dan abstrak artikel ini, kami menggunakan perangkat lunak VosViewer untuk membuat jaringan bibliometrik (Gambar 1 ). Jaringan ini mewakili istilah yang paling sering muncul, dikelompokkan berdasarkan warna untuk menunjukkan kedekatannya dengan area minat lain di lapangan. Analisis kami mengungkapkan tiga kelompok utama berikut: istilah yang terkait dengan fungsi mitokondria dan apoptosis (merah), polifenol dan obesitas (hijau), dan penyakit degeneratif lainnya dan pencegahannya (biru).
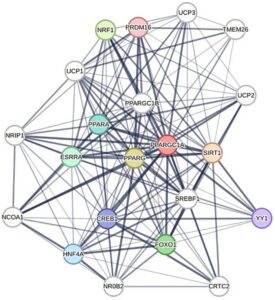
3 STRING Jaringan Interaksi Protein PGC-1α yang Dihasilkan
String ( https://string-db.org/ ) berfungsi sebagai sumber daya untuk menyelidiki interaksi protein-protein, mengintegrasikan asosiasi yang ditentukan secara eksperimental dan diprediksi secara komputasi untuk menghasilkan jaringan asosiasi fungsional. Di bawah ini, kisi asosiasi dihasilkan dengan tingkat pertama interaksi protein PGC-1α (Gambar 2 ) untuk manusia. Seperti yang dapat kita lihat dalam jaringan ini, data yang tersedia mengkorelasikan PPARGC1α (gen yang mengkodifikasi PGC1α); PPARα (reseptor yang diaktifkan oleh proliferator peroksisom alfa); PPARG (reseptor yang diaktifkan oleh proliferator peroksisom gamma), dan SIRT1 (Sirtuin 1) adalah protein paling umum yang terkait dengan PGC-1α dalam fungsi, bekerja sama erat untuk mempertahankan fungsi mitokondria dan energetik tubuh. Di antara tingkat kedua (lingkaran putih), isoform UCP dan SREBF1 (faktor transkripsi pengikatan elemen pengatur steroid 1) adalah yang paling sering dikutip dalam studi polifenol. Meningkatnya kadar UCP dalam sel-sel adiposa dikaitkan dengan “pencoklatan”, suatu aspek yang didapat oleh jaringan adiposa dan jaringan adiposa coklat karena sejumlah besar mitokondria yang meningkat dalam jumlah, ukuran, dan biasanya lamela.
4 Meningkatkan Pencoklatan Jaringan Adiposa Putih dan Aktivasi Jaringan Adiposa Coklat
Aktivasi BAT dan pencoklatan WAT adalah proses multifaset, dipicu oleh beberapa jalur metabolisme. Kompleksitas intrinsik jalur seluler, yang dicirikan oleh sensitivitas dan responsivitas tinggi terhadap beberapa rangsangan, menghalangi penggambaran jalur pensinyalan tunggal utama. Dengan mengingat hal ini, PGC1α sering disebut sebagai “pengatur utama proses pencoklatan,” yang berfungsi sebagai reseptor sentral dari banyak sinyal eksternal dan internal. Protein ini diketahui berinteraksi dengan setidaknya 162 molekul [ 18 ], termasuk banyak protein kecil hilir dan hulu. Meskipun PGC-1α tidak sepenuhnya diperlukan agar pencoklatan terjadi, sel yang kekurangan PGC-1α atau isoform PGC-1β menunjukkan gangguan biogenesis mitokondria dan berkurangnya ekspresi sebagian besar protein terkait [ 19 ]. Interaksi protein yang mendukung biogenesis mitokondria, peningkatan metabolisme lipid, dan pengaktifan termogenesis non-menggigil dijelaskan di bawah ini dan diilustrasikan dalam Gambar 3 .
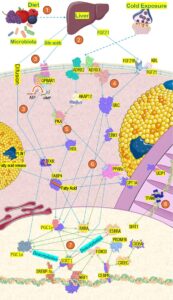
Termogenesis dimulai secara fisiologis oleh sensasi dingin, olahraga, atau asupan makanan [ 20 , 21 ], yang merangsang pelepasan norepinefrin dari saraf simpatik untuk mengaktifkan reseptor adrenergik (ADBR) di jaringan adiposa [ 22 ]. Selain itu, AMPKα2 diketahui meningkatkan ekspresi UCP1 melalui PGC1α untuk meningkatkan pengaktifan jaringan adiposa putih (WAT) dan jaringan adiposa coklat (BAT) [ 23 , 24 ].
5 Mencapai Aktivasi PGC1α dengan Konsumsi Polifenol
5.1 Konsumsi Oral
Polifenol mengalami transformasi metabolik substansial selama pencernaan dan pensinyalan intraseluler berikutnya sebelum memberikan efek termogeniknya pada jaringan adiposa. Efek ini terutama dikaitkan dengan molekul yang lebih kecil, seperti asam kafeat dan protokatekuk, asam empedu (BA), dan asam lemak rantai pendek (SCFA), yang berasal dari biotransformasi yang dimediasi oleh enzim pencernaan dan mikrobiota usus (GM) [ 12 ]. GM memainkan peran penting dalam proses ini, khususnya bakteri, sebagaimana dibuktikan oleh pelemahan termogenesis yang diinduksi ekstrak polifenol setelah pemberian antibiotik [ 25 ]. Molekul yang lebih kecil ini mencapai hati dan meningkatkan pelepasan setidaknya faktor pertumbuhan fibroblas 15 dan 21 (FGF15/21) [ 26 , 27 ] dan asam empedu [ 28 ], yang kemudian bersirkulasi dalam aliran darah dan mencapai jaringan lain.
FGF21 mengikat reseptor spesifiknya pada permukaan sel adiposa (FGF21R), fusi yang juga memerlukan β-klotho (KLB) [ 29 ]. Ini memulai kaskade pensinyalan kompleks yang mengarah pada peningkatan metabolisme lipid dan stimulasi termogenesis [ 30 ]. FGF21R juga bekerja bersama dengan β-adrenergik tipe 3 (ADRB3) untuk menginduksi termogenesis, karena tikus FGF21-KO tidak responsif terhadap pencoklatan melalui agonis adrenergik [ 31 ]. Di sisi lain, FGF21 tidak penting untuk pencoklatan jaringan adiposa putih inguinal (iWAT) pada tikus yang terpapar dingin [ 32 ], dan produksi FGF21 yang dirangsang oleh konsumsi fruktosa tampaknya tidak mengaktifkan termogenesis pada wanita Jepang saja [ 33 ]. Menariknya, pada manusia, ADBR3 yang ditemukan pada adiposit relatif lebih sedikit [ 34 ]. Bukti kuat menghubungkan stimulasi termogenesis manusia oleh agonis noradrenergik dengan isoform ADBR2, bukan ADBR3, yang terakhir secara tradisional dikaitkan dengan respons noradrenergik pada tikus dan mencit [ 35 ]. Ini adalah perbedaan penting yang harus diingat oleh para peneliti termogenesis ketika mengevaluasi stimulasi β-adrenergik dan termogenesis pada hewan pengerat.
Sebaliknya, FGF21 tampaknya diperlukan agar senyawa fenolik dapat memberikan efek menguntungkan. Dalam intervensi dengan kurkumin dan resveratrol, tikus jantan yang kekurangan FGF21 (FGF21-KO) tidak menunjukkan penurunan yang sama dalam total gliserida dan perbaikan dalam toleransi lemak seperti yang memiliki fungsi FGF21 yang utuh [ 31 ]. Namun demikian, untuk tikus obesitas yang diinduksi oleh diet, FGF21 memainkan peran penting dalam menjaga homeostasis tubuh dalam kondisi dingin sedang [ 36 ], yang menyiratkan bahwa FGF21 mungkin lebih terlibat dalam metabolisme lipid dan kalsium, dan hanya secara tidak langsung terlibat dengan bahan bakar termogenesis UCP1. Karena adiposit juga dapat menghasilkan faktor ini dalam mode autokrin [ 37 , 38 ], peran FGF21 mungkin cocok dengan amplifikasi sinyal, di mana adiposit merangsang seluruh jaringan untuk menginduksi pencoklatan sebagai respons terhadap pensinyalan awal hati [ 30 , 39 ].
Dari perspektif asam empedu, BA primer yang disintesis di hati dikonjugasikan dengan taurin pada tikus dan glisin pada manusia [ 40 ]. Hidrolase garam mendekonjugasikan asam amino ini dari BA primer, yang memungkinkan terjadinya biotransformasi bakteri. Proses ini melibatkan reaksi seperti 7α/β-dehidroksilasi, dehidrogenasi, dan epimerisasi, yang menghasilkan produksi BA sekunder [ 41 ]. Induksi signifikan dari subfamili kaset pengikat ATP anggota G 5/8 (ABCG5/G8), pengangkut asam empedu, diamati pada sel hati dan enterosit yang diobati dengan polifenol [ 42 – 44 ], tetapi peran dan kelimpahan famili ini pada adiposit yang terpapar polifenol masih kurang dijelaskan. BA dipelajari lebih baik dengan mengaktifkan reseptor asam empedu berpasangan protein G 1 (GPBAR1) di membran sel, dan ekspresinya diatur oleh agen polifenol [ 45 , 46 ]. GPBAR1 pada gilirannya merangsang produksi cAMP, pembawa pesan kedua yang terlibat dalam banyak peran seluler [ 47 , 48 ]. Peningkatan konsentrasi cAMP ini diketahui mengaktifkan protein kinase A (PKA), yang pada gilirannya memfosforilasi lipase sensitif hormon (HSL) dan perilipin 1 (PLIM1), yang mengakibatkan pemecahan trigliserida yang disimpan dalam jaringan adiposa [ 16 , 49 ]. cAMP juga mengaktifkan protein pengikat elemen respons cAMP (CREB), yang berinteraksi dengan PGC-1α untuk mendorong pencoklatan [ 50 ]. Antosianin mampu meningkatkan fosforilasi CREB melalui aktivasi PKA, yang menginduksi peningkatan regulasi yang dimediasi CREB [ 51 ].
5.2 Sinyal Intraseluler Dari Reseptor FXR ke PGC1α
Reseptor farnesoid X (FXR) memiliki peran kompleks dalam aktivasi adiposit coklat, bertindak sebagai agonis dan antagonis [ 25 , 40 , 52 ]. Agonis FXR, seperti fexaramine, telah terbukti meningkatkan ekspresi gen browning dalam jaringan adiposa coklat (BAT) dan jaringan adiposa putih gonadal (gWAT) [ 53 ]. Suplementasi polifenol pada tikus obesitas diabetes (db/db) dapat meningkatkan metabolisme glukosa dengan menghambat FXR, yang mengarah pada peningkatan sintesis asam empedu primer dan penurunan kadar asam empedu sekunder, pengurangan ini semakin melemahkan aksi FXR, karena mereka bertindak sebagai agonis FXR [ 40 ]. Mekanisme di mana asam empedu hidrofobik memasuki sel-sel ini dan mengikat FXR masih harus diklarifikasi sepenuhnya, dengan transpor aktif dan difusi pasif menjadi kemungkinan potensial.
FXR dapat membentuk heterodimer dengan reseptor retinoid X α (RXRA), yang secara langsung mengikat PGC1α dan memodulasi aktivitasnya [ 40 ]. PGC1α mengirimkan sinyal ke PR domain containing 16 (PRDM16), sebuah protein yang memainkan peran penting dalam membedakan sel menjadi sel lemak coklat [ 54 ]. Ia bekerja bersama dengan CCAAT-enhancer-binding protein β (C/EBP-β), untuk memicu peralihan ini, karena memaksa ekspresi PRDM16 dan C/EBP-β pada sel kulit normal (fibroblast) sudah cukup untuk mengubahnya menjadi sel lemak coklat [ 19 ]. PRDM16 adalah jenis protein zinc finger, yang diketahui berpartisipasi langsung di daerah pengikatan DNA yang memungkinkan transkripsi gen terkait pencoklatan [ 55 ]. PRDM16-C/EBPβ, PGC1α, dan PGC1β semuanya mungkin ditekan oleh peningkatan ekspresi FXR (Yang et al., 2023b). SMYD1 (histone-lysine N-methyltransferase) baru-baru ini dikaitkan dengan termogenesis yang diinduksi dingin atau diet, menunjukkan bahwa SMYD1 diaktifkan dalam WAT dan BAT, bekerja bersama faktor transkripsi lainnya (Cicatiello et al., 2024) [ 56 ]. Ekspresi protein ini lebih tinggi pada adiposit krem dibandingkan pada adiposit putih, dan pembungkamannya menyebabkan gangguan aktivitas mitokondria.
Sirtulin 1 (SIRT1), protein nuklir, juga ditemukan di mitokondria [ 57 ], berinteraksi dengan dan mendeasetilasi PGC-1α dalam cara yang bergantung pada NAD+, dan interaksi ini memengaruhi ekspresi gen glukoneogenik yang dimediasi oleh PGC-1α [ 58 , 59 ]. Namun, SIRT1 mungkin tidak mengatur efek PGC-1α pada gen mitokondria [ 60 ]. Mengingat penginderaan fluktuasi nutrisi yang bergantung pada NAD+, SIRT1 kemungkinan berfungsi sebagai pengatur utama ekspresi gen yang bergantung pada PGC-1α [ 58 ].
5.3 Penghambatan Kinase Keluarga Src
Kinase famili Src (SRC) merupakan rem pengatur pada jaringan adiposa cokelat dan krem yang sedang berkembang. Pada manusia, ditemukan bahwa SRC berkorelasi positif dengan diferensiasi adiposit [ 61 ]. Penghambatan SRC melalui berbagai pendekatan telah terbukti mendorong diferensiasi preadiposit cokelat dan pencoklatan preadiposit putih yang mengatur protein terkait termasuk PGC-1α [ 62 ]. Pengamatan penting adalah penurunan aktivitas SRC selama pematangan adiposit cokelat dan putih, sementara SRC diekspresikan pada tingkat yang jauh lebih tinggi di WAT daripada di BAT [ 62 ]. SRC diaktifkan oleh ADBR3, dan diperlukan untuk mengaktifkan kinase yang diatur sinyal ekstraseluler (ERK1/2) dan mencapai kofaktor nuklir di adiposit cokelat [ 63 , 64 ].
Reseptor gamma yang diaktifkan oleh proliferator peroksisom (PPARG), koaktivator PGC-1α lainnya, terbukti berinteraksi langsung dengan SRC, memengaruhi aktivitas masing-masing. SRC dapat menekan aktivitas transkripsi PPARG dengan cara yang tidak bergantung pada aktivitas kinase melalui interaksi protein-protein [ 65 ] dan melalui ERK1/2 [ 66 , 67 ]. Selain itu, SRC berinteraksi dengan PPARG untuk mengurangi lipolisis melalui protein pengikat asam lemak 4 (FABP4) [ 65 ].
Beberapa molekul polifenol telah diidentifikasi sebagai penghambat SRC, dengan implikasi potensial terhadap peradangan dan karsinogenesis [ 68 , 69 ]. Meskipun peran SRC menjanjikan dalam regulasi adipogenesis dan termogenesis, penelitian yang secara khusus meneliti interaksi langsungnya dengan polifenol dalam konteks ini terbatas.
5.4 Reseptor Alfa Terkait Estrogen (ERRα)
ERRα diketahui terlibat dalam termogenesis dan aktivasi selektif protein dapat menginduksi kelahiran adiposit [ 70 ]. ERRα telah lama diketahui berinteraksi secara fisik dengan PGC1α setelah fosforilasi [ 71 , 72 ]. Bersama-sama, mereka mengatur ekspresi gen yang terkait dengan proses metabolisme, fungsi mitokondria, dan aktivitas transkripsi [ 73 ]. Selain itu, ERRα membentuk heterodimer dengan PGC1α yang mengikat elemen respons di wilayah promotor gen ESRRA, yang menyebabkan peningkatan ekspresinya [ 74 ].
Kaskade sinyal estrogen-Wnt dapat meningkatkan ekspresi FGF21 dalam jalur yang independen dari PPARα dan PGC1α [ 75 ]. Banyak zat polifenol, yang dikenal sebagai fitoestrogen, dapat berinteraksi dengan ERRα dan mengatur metabolisme energi [ 76 , 77 ]. Interaksi dengan hormon wanita dan jalur lipid/termogenik ini mungkin menjelaskan sebagian mengapa pria, tetapi tidak wanita, menunjukkan gangguan toleransi lemak dalam beberapa penelitian [ 37 , 78 ].
5.5 Ko-Aktivator Transkripsi Lainnya
Di antara protein lain yang telah dipelajari dengan baik yang dimodulasi oleh aktivitas polifenol, Nuclear Respiratory Factor 1 (NRF1), Sterol Regulatory Element Binding Protein 1-c (SREBP-1c), dan mitochondrial transcription factor A (TFAM) adalah protein yang terkait erat dengan koaktivator PGC1 dan memodulasi aktivitas transkripsi banyak gen termogenik [ 79 – 81 ]. NRF1 terlibat dengan gen metabolik utama yang mengatur gen nuklir yang diperlukan untuk respirasi mitokondria dan transkripsi dan replikasi DNA mitokondria [ 82 ]. SREBP-1c adalah protein utama yang mengatur lipogenesis [ 83 ], yang diketahui berinteraksi langsung dengan SIRT1 [ 84 ]. TFAM mengikat langsung dalam mtDNA dan terlibat dengan replikasi, perbaikan, dan biogenesis mitokondria [ 85 ].
5.6 Karnitin Palmitoiltransferase
Karnitin palmitoiltransferase 1A (CPT1A) adalah enzim penting yang ditemukan di membran mitokondria luar yang memainkan peran vital dalam metabolisme asam lemak, termasuk transportasi asam lemak, masuknya mitokondria, dan produksi energi berikutnya melalui pengangkutan karnitin dengan karnitin palmitoiltransferase 2 (CPT2) di membran dalam [ 86 ]. Knock-out PGC1A dapat menurunkan tingkat transkripsi kedua protein [ 87 ] dan kelimpahannya diatur naik oleh pemberian polifenol dalam sel manusia [ 88 , 89 ] dan tikus [ 90 ]. Bersama-sama, mereka bertanggung jawab untuk memfasilitasi masuknya lipid ke dalam mitokondria, memicu pelepasan yang dimediasi UCP1 dan karenanya penting untuk termogenesis.
6 Senyawa Fenolik dalam Model Sel
Meskipun terdapat banyak jenis senyawa fenolik, mayoritas senyawa yang diuji diketahui memodulasi lipogenesis dan obesitas sampai batas tertentu, juga mendorong biogenesis mitokondria (Tabel 1 ). Perlakuan quercetin secara signifikan meningkatkan ekspresi UCP1, PGC1α, dan Tfam pada adiposit 3T3-L1, yang menunjukkan potensinya untuk menginduksi pencoklatan pada sel manusia [ 91 ]. Efek quercetin pada induksi UCP1 tampaknya dimediasi oleh interaksinya dengan PPARγ sebagaimana dibuktikan oleh ekspresi UCP1 diferensial yang diamati pada adiposit 3T3-L1 yang diobati dengan quercetin dengan atau tanpa antagonis PPARγ [ 91 ]. Juga dengan adiposit 3T3-L1, diamati bahwa ekstrak Smilax china L., engelitin, quercetin, dan asam caffeic dapat mengubah sel lemak putih menjadi sel lemak seperti krem atau coklat [ 92 ]. Banyak enzim yang terlibat dalam regulasi lipolisis (lipase trigliserida adiposa [ATGL] dan PKA) dan β-oksidasi (PPARα, CPT-1, oksidase asam karboksilat [ACO]) yang mendorong pemecahan asam lemak dan pengeluaran energi. Lebih jauh lagi, ekstrak dan monomernya secara ampuh merangsang program pencoklatan dengan meningkatkan gen dan protein spesifik cokelat seperti UCP1, PGC-1α, dan PRDM16, bersama dengan penanda spesifik krem [ 92 ]. Sebuah studi menggunakan ekstrak buah honeysuckle biru mencatat pengurangan akumulasi lemak dengan menurunkan kadar protein yang terlibat dalam produksi lemak dan mengaktifkan enzim yang membakar lemak, juga meningkatkan penanda untuk lemak krem dalam cara yang bergantung pada dosis, termasuk UCP1, PPARγ, dan C/EBPα [ 93 ].
| Honeysuckle biru ( Lonicera caerulea ) | Ekstrak fenolik | Sel 3T3-L1 | 50 dan 100 µg/mL | 6 hari | ⇑ ACC, fosforilasi AMPK, Cd137, Tmem26, UCP1 (PX)
∅ TG, LIPE, ATGL ⇓ Akumulasi lipid pada adiposit, FAS, SREBP-1c, PPARγ, C/EBPα (PX) |
[ 93 ] |
| Obat Resveratrol | Stilbenoid | adiposit hMADS | 0,1–10 µm/ml | 4 hari | ⇑ UCP1 hingga 1 µm (PX, GX), SIRT, CIDEA (GX)
⇓ UCP1 > 1 µm (PX, GX), CIDEA (GX) |
[ 117 ] |
| 6-Gingerol | jahe | Preadiposit 3T3-L1 | 20-150 µg/ml | 6 hari | ⇑ UCP1, PRDM16, PGC1α, PPARγ, C/EBPα, AP2, Cidea, Cited1, SIRT1, Tmem26 (GX)
⇓ TG |
[ 94 ] |
| Smilax china L. engeltin; quercetin; asam kafeat | Glukosida fenolik, flavonoid, asam fenolik | adiposit 3T3-L1 | 40 µg/ml | 24 jam | ⇑ ATGL, UCP-1, CPT, ACO, PPARα, PGC-1α, PRDM16 PGC-1α, PRDM16, Nrf1 dan Tfam, Tmem26, Tbx1, CD137, β3-AR, PKA dan pAMPKα (GX dan PX).
∅ Viabilitas sel ⇓ Kandungan lipid |
[ 92 ] |
Ekstraksi etil asetat dari 6-gingerol, salah satu komponen utama jahe, mampu menginduksi diferensiasi adiposit coklat dari preadiposit 3T3-L1, namun tidak seperti ekstraksi air dari akar yang sama, meningkatkan ekspresi UCP1, Prdm16, dan PGC1α, juga menggandakan ekspresi NRF1 dan TFAM [ 94 ].
7. Pengaturan Termogenesis oleh Senyawa Fenolik pada Model Hewan Pengerat
Dari Tabel 2 , kita dapat mengamati bahwa UCP1, PCG1α, PRDM16, CIDEA, SIRT1, PPARγ, dan CPT1A adalah protein yang paling umum mengalami peningkatan regulasi di banyak jaringan setelah pemberian polifenol. Mengenai studi zat murni, dalam sebuah studi dengan resveratrol dan pterostilbene, diamati bahwa keduanya dapat mencegah akumulasi trigliserida dalam jaringan adiposa coklat dan meningkatkan ekspresi UCP1, dengan resveratrol juga meningkatkan ekspresi SIRT3, sementara tidak ada senyawa yang secara signifikan mempengaruhi aktivasi PGC-1α dalam dosis (30 mg/kg) yang diberikan; hasil ini menunjukkan bahwa efeknya pada kapasitas termogenik bergantung pada status metabolik dan kondisi spesifik dari diet tinggi lemak, tinggi fruktosa [ 95 ]. Dalam studi lain, pemberian pterostilbene diamati secara signifikan meningkatkan kadar protein UCP1, meskipun tidak ada perubahan yang diamati dalam ekspresi mRNA UCP1 [ 96 ]. Tikus yang diberi diet tinggi lemak yang dilengkapi dengan dosis asam protocatechuic yang dapat dicapai melalui diet (0,003%) dapat mengalami pengurangan yang signifikan dalam penambahan berat badan, meningkatkan sensitivitas insulin, dan mengurangi akumulasi lemak di hati [ 97 ]. Efek-efek ini tidak terkait dengan peningkatan pembakaran lemak di jaringan adiposa, tetapi lebih cenderung pada peningkatan pemrosesan lemak oleh PCA di dalam hati dan jaringan adiposa melalui peningkatan aktivitas enzim CPT1A [ 97 ].
| Senyawa fenolik | Kelas | Model | Dosis harian | Waktu | Temuan | Referensi |
|---|---|---|---|---|---|---|
| Magnolol, Hinokiol | Lignan | Mouse | 30mg/kg | 8 minggu | ⇑ PPARα, PPARγ dan UCP1, oksidasi asam lemak, peningkatan ekspresi dan sekresi FGF21, pencoklatan jaringan
⇓ lipogenesis |
[ 100 ] |
| Asam protocatechuic | Asam fenolik | Mouse | 0,003% | 16 minggu | ⇑ Sensitivitas insulin
∅ Tidak ada perubahan UCP1 pada jaringan ⇓ Peningkatan berat badan dan berkurangnya steatosis hati. |
[ 97 ] |
| Pterostilbene dan resveratrol | Stilbenoid | Mouse | 30mg/kg | 8 minggu | ⇑ UCP1 dan SIRT3 (PX) untuk resveratrol
∅ Tidak ada perubahan pada asupan makanan, berat iBat, ekspresi SIRT1, ATGL, CD36, FATP1, NRG4, EPDR1, GLUT4, dan ekspresi HSL ⇓ BW untuk resveratrol, akumulasi trigliserida dalam ibat, mencegah aktivitas CPT-1A yang lebih besar, ekspresi NRF1 untuk stilbene, ekspresi FAS untuk keduanya |
[ 95 ] |
| Anggur konvensional dan anggur organik | Polifenol | Mouse | 100mg/kg | 10 minggu
(6,12,18 fotoperiode) |
⇑ Ekspresi UCP1 dan leptin pada BAT, pencoklatan iWAT, pengeluaran energi
∅ TG, TC |
[ 106 ] |
| Rimpang Smilax china L. | Ekstrak | Mouse | 100–400mg/kg | 10 minggu | ⇑ GLUT4, IRS1, IRS2, AKT, ACO, PKA (GX); AKT, p-AKT, GLUT4, UCP-1, AMPK, dan p-AMPK (PX) di hati
∅ Asupan Makanan ⇓ BW, (SL) HDL LDL, TC, dan TG, GLU, Insulin, IR, IL-6, MCP-1, dan TNF-α, akumulasi lemak; mTORC1, SREBP1c, FAS, HMGCR, FOXO1 (GX); SREBP1c, FAS, HMGCR, p-IKBα, dan P65 (PX) di hati |
[ 99 ] |
| Kurkumin | Kurkuminoid | Mouse | 20mg/kg | 6 minggu | ⇑ Ekspresi UCP1, PRDM16 di BAT dan iWAT (PX), GT
∅ BB, adipositas, asupan makanan, berat iBAT, PGC1-α (PX) ⇓ TC, TG, glukosa, insulin |
[ 98 ] |
| Tetrahidrokurkumin | Kurkuminoid | Mouse | 20 dan 100 mg/kg | 14 minggu | ⇑ UCP1, Produksi panas untuk 20 mg/kg, adiponektin (serum dan WAT), GLUT4 dalam adiposit, CPT-1, PPARα (PX) di hati
∅ Hati, ginjal, dan pankreas, Berat kelelawar ⇓ Glukosa darah, berat limpa, TG, TC, asupan makanan 100 mg/kg, infiltrasi makrofag dan modulasi M1/M2 pada eWAT, steatosis hati, GOT, GTP, ACC, FAS (PX) di hati |
[ 118 ] |
| Ekstrak kakao | flavanol | Mouse | 50mg/kg | 2 minggu | ⇑ Ekskresi ND, AD, UCP-1, PGC-1α, dan PRDM16 (GX) di BAT, PGC-1α, CD137, TMEM26; PRDM16 (GX) di iWAT
∅ UCP-1 (PX); PRDM16, TMEm26, TBX-1 (GX) di BAT; UCP-1, PGC-1α, CD137, TMEM26, dan TBX-1 (GX) di iWAT |
[ 104 ] |
| Ekstrak blueberry dan cranberry | Ekstrak fenolik | Mouse | Ekstrak 1% dan 2% | 24 minggu | ⇑ SCFA tinja
∅ Asupan energi, MCP-1 dan IL-1β dalam plasma ⇓ BW, WAT, Berat Hati, TC, TG, LPS, TNF-α dalam plasma, SFA, MUFA, PUFA dalam hati, rasio F/B dalam cranberry 2%, |
[ 107 ] |
| Kuersetin | Flavonoid | Tikus C57BL/6 | 0,05% | 9 minggu | ⇑ UCP1, PPARγ dan PGC1α, TFAM, NRF-1, PKA, PRDM16, CIDEA, TMEM26 (PX), fosforilasi AMPK di WAT, NE dalam plasma, β3AR (GX) di WAT, UCP1 (PX) di BAT
∅ SELESAI ⇓ Berat badan WAT, kenaikan berat badan |
[ 91 ] |
| Teh hijau | Ekstrak fenolik | Tikus C57BL/6 | 500mg/kg | 12 minggu | ⇑ Sensitivitas insulin, UCP1, Xbp1s (GX) di BAT, PPARα, CPT1b, Adrb3, Irf4, PPARGC1α, PPARΓ, CIDEA, PDE1B, UCP1, LXR (GX) SWAT
∅ FI, PPARΑ, CPT, ADRB3, IRF4, PPARGC1A, PPARG, CIDEA, DIO2, PDE1B (GX) DI KELALAWAR ⇓ Peningkatan berat badan, eWAT dan sWAT wt, Glukosa, Insulin dalam plasma, IL-1β, IL-6, TNF-α, IL17, IFN-γ, dan MCP-1 dalam plasma, HSPA5 (GX) dalam BAT dan WAT, |
[ 101 ] |
| preadiposit 3T3-F442A | konsentrasi 0,19% | 8 hari | ⇑ Konsumsi oksigen, PPARα, Adrb3, PPARGC1α, CIDEA, dan UCP1 (GX) | |||
| Ekstrak polifenol cranberry | Ekstrak fenolik, antosianin | Tikus C57BL/6J | 63,8 mg/kg | 16 minggu | ⇑ Sensitivitas insulin, VO 2 , EE, ATGL, CPT1A, MCAD (GX) di hati, CPT1A, MCAD, UCP1, PGC1α, CIDEA, Cd137, TBX1 (GX) di iWAT, PPARA, MCAD, UCP1, PGC1α, CIDEA, PRDM16 (GX) di BAT; TMEM26, HOXC8, CD137 di eWAT
∅ FI, Bat wt, VCO 2 , CPT1A, MCAD, PPARA, UCP1 (GX) dalam eWAT ⇓ Peningkatan berat badan, Berat hati, TNFA, MCP1 (GX) di hati |
[ 102 ] |
| Apel | Polifenol apel | Tikus C57BL/6J | 5% polifenol apel | 4 minggu | ⇑ OXPHOS, CD137 (PX) dalam eWAT; UCP1, CIDEA, TBX1, CD137, FGF21 (GX), UCP1, OXPHOS, PGC1 α, FGF21 (PX), ND dalam iWAT
∅ Asupan energi, UCP1, CIDEA, NDUFB8, SDHB, UQCRC2, COX I, dan ATP5A (GX) di BAT; UCP1, CIDEA, TBX1, CD137 (PX) PGC1 α di BAT dan eWAT dan (GX) di eWAT ⇓ Keuntungan BW, BAT, eWAT, iWAT wt, |
[ 103 ] |
| Sel induk yang berasal dari jaringan lemak (ASC) | 5–100 µg/ml | 4 hari | ∅ UCP1, PGC-1α, FGF21 (GX) dan (PX) OXPHOS (PX), diferensiasi adiposit krem.
⇓ Fabp4 (GX) |
|||
| Pterostilbene | Stilbenoid | Tikus C57BL/6 | 22, 45, dan 90 mg/kg/hari | 30 minggu | ⇑ GT, CIDEA, EBF2, PGC1α, PPARγ, Sirt1, dan Tbx1(GX) UCP1 (PX)
∅ CITED1, FGF21, PAT2, PPARα, UCP1 (GX) ⇓ BW |
[ 96 ] |
| Preadiposit 3T3-L1 | 5 mikron | 1 hari | ⇑ CIDEA, FGF21 (GX), UCP1 (PX)
∅ CITED1, EBF2, PAT2, PGC-1α, PRDM16, SIRT1, TBX1, UCP1 (GX) ⇓ PPARγ (GX) |
|||
| asam lemak; | Stilbenoid | Tikus C57BL/6 | 0,5% | 12 minggu | ⇑ UCP1, PRDM16, SIRT1, PGC-1α, PPARα, adipo (PX)
∅ mWAT berat, ⇓ Berat Badan, LDL, gWAT dan rWAT berat badan, |
[ 119 ] |
| Oksifera | 0,1% dan 0,5% | ⇑ PRDM16, SIRT1, PGC-1α, C/EBPβ (PX)
∅ MES berat, gWAT berat; UCP1, CPT1, adipo(PX) ⇓ Berat jenis, rWat; |
||||
| Ekstrak blueberry | Tikus | ⇑ UCP1, PRDM16, PCG1α, PPARα, DIO2, COX7a1, COX8b, CIDEA, Errα (GX) UCP1, PCG1α (PX),
∅ ⇓ TG, LEP, |
[ 25 ] | |||
| Senyawa 18a | flavonoid | Tikus C57BL/6J | 20–40mg/kg | 9 minggu | ⇑ UCP1, PRDM16, PCG1α, TGR5, DIO2 (GX) UCP1, PRDM16, PCG1α (PX), suhu rektal
∅ ⇓ iBAT, iWAT, eWAT berat, BW, GT |
[ 120 ] |
Singkatan : ACC, asetil-CoA karboksilase; ACO, asil-CoA oksidase; AD, adrenalin; Adipo, adiponektin; Adrb3, adrenoseptor beta 3; AKT, protein kinase B; AMPKα, subunit alfa protein kinase terfosforilasi teraktivasi; AP2, protein adaptor 2; ATGL, lipase trigliserida adiposa; ATP5A, subunit Kompleks V; C/EBP α, CCAAT/protein pengikat penambah alfa; C/EBPβ, CCAAT/protein pengikat penambah beta; CD36, gugus diferensiasi 36; CD137, gugus diferensiasi 137; CIDEA, subunit alfa faktor fragmentasi DNA pemicu kematian sel; COX I, Kompleks IV; CPT, karnitin palmitoiltransferase 1ª; CPT-1A, karnitin palmitoiltransferase 1ª; Dio2, Iodothyronine deiodinase 2; EE, pengeluaran energi; EPDR1, Protein terkait ependim 1; FAS, sintase asam lemak; FATP1, Protein pengangkut asam lemak 1; FI, asupan makanan; FOXO1, Protein kotak Forkhead O1; GLUT4, pengangkut glukosa Tipe 4; GOT, transaminase glutamat-oksaloasetat; GPT, transaminase glutamat-piruvat; GT, toleransi glukosa; GX, ekspresi gen; gWAT, jaringan adiposa putih gonad; HMGCR, 3-hidroksi-3-metilglutaril-koenzim A reduktase; HOXC8, protein C8 yang mengandung homeobox; HSL, lipase peka hormon; HSPA5, protein syok panas Keluarga A (Hsp70) Anggota 5; iBAT, berat jaringan adiposa coklat interskapular; IL-1β, interleukin-1 beta; IR, resistensi insulin; Irf4, Faktor pengatur interferon 4; IRS1, substrat reseptor insulin 1; IRS2, substrat reseptor insulin 2; LIPE, lipase peka hormon; MCAD, dehidrogenase asil-CoA rantai menengah; MCP-1, Protein-1 kemoatraktan monosit; mTORC1, kompleks target rapamycin mamalia 1; MUFA, asam lemak tak jenuh tunggal; mWAT, jaringan adiposa putih mesenterika; NE, norepinefrin; NRF1, Faktor pernapasan nuklir 1; NRG4, Neuregulin 4; NDUFB8, subunit Kompleks I; Pde1b, fosfodiesterase 1B; PGC-1α, koaktivator gamma reseptor proliferator peroksisom 1 alfa; PKA, protein kinase A; PPARα, reseptor proliferator peroksisom alfa; PPARγ, reseptor proliferator peroksisom gamma; PRDM16, protein yang mengandung domain PR 16; PX, ekspresi protein; PUFA, asam lemak tak jenuh ganda; rWAT, jaringan adiposa putih retroperitoneal; SDHB, subunit Kompleks II; SCFA, asam lemak rantai pendek; sWAT, jaringan adiposa putih subkutan; SFA, asam lemak jenuh; SIRT1, Sirtuin 1; SREBP1c, protein pengikat elemen pengatur sterol 1c; TBX1, faktor transkripsi kotak-T 1; TC, kolesterol total; TG, trigliserida serum; Tfam, faktor transkripsi A, mitokondria; Tmen26, protein transmembran 26; UCP-1, protein pelepas 1; UQCRC2, subunit Kompleks III; VCO2, produksi karbon dioksida maksimum; VO2, konsumsi oksigen maksimum; Xbp1s, protein pengikat kotak-X 1 yang disambung; wt, berat.
Kurkumin yang diberikan kepada tikus obesitas hamil dapat meningkatkan metabolisme keturunan jantan dewasa. Penelitian terbaru menunjukkan bahwa mereka bertambah lebih sedikit berat badan, memiliki kontrol gula darah yang lebih baik, dan menurunkan kolesterol, yang tampaknya terkait dengan peningkatan gen untuk pembakaran lemak cokelat dan krem, senyawa tersebut dapat membantu keturunan menghindari beberapa efek negatif dari diet tinggi lemak ibu [ 98 ]. Metabolit kurkumin, tetrahidrokurkumin, testis pada tikus obesitas diabetes yang diberikan dalam dosis 20 dan 100 mg/kg memperbaiki hiperglikemia, tetapi mekanismenya berbeda. 20 mg/kg meningkatkan sinyal adiponektin (AdipoR1/R2, APPL1) yang mengarah pada peningkatan sensitivitas insulin, metabolisme glukosa, dan fungsi sel beta. Selain itu, meningkatkan pengeluaran energi melalui pencoklatan WAT dan peningkatan regulasi UCP1. Dosis 100 mg/kg menunjukkan asupan makanan yang berkurang dan hasil positif yang kurang ekspresif, tetapi ekspresi UCP1 yang lebih baik.
Dalam studi yang bergantung dosis, pemberian ekstrak rimpang S. china L. meningkatkan ekspresi AKT, p-AKT, GLUT4, UCP-1, AMPK, dan p-AMPK di hati, dan menekan ekspresi SREBP1c [ 99 ], yang menunjukkan bahwa polifenol secara signifikan meningkatkan ekspresi PGC-1α, mengaktifkannya melalui jalur pensinyalan substrat reseptor insulin/protein kinase B (IRS/AKT), AMPK, dan faktor nuklir kappa-light-chain-enhancer of activated B cells (NF-κB). Efek polifenol S. china L. pada parameter obesitas pada dosis rendah (100 mg/kg), sedang (200 mg/kg), dan tinggi (400 mg/kg) secara signifikan mengurangi penambahan berat badan dan indeks jaringan adiposa intraperitoneal, dengan efek yang paling menonjol diamati pada dosis tinggi. Selain itu, dosis yang lebih tinggi berhubungan dengan penurunan yang lebih besar pada kadar lipid serum dan faktor inflamasi, yang meningkatkan sensitivitas insulin. Magnolol dan Honokiol, senyawa yang ditemukan di Magnolia officinalis , dilaporkan berfungsi sebagai agonis ganda reseptor PPARα dan PPARγ, dalam mode yang bergantung pada dosis. Percobaan pada tikus jantan (30 mg/kg selama 8 minggu) dengan kedua senyawa tersebut meningkatkan produksi FGF21 di hati, serta kadar CPT1A, CPT1B, dan CPT2, meningkatkan pencoklatan jaringan dan termogenesis sekaligus mengurangi berat jaringan adiposa [ 100 ].
Pengobatan dengan teh hijau 500 mg/kg yang diberikan lewat mulut selama 12 minggu mampu secara efektif mengurangi asam lemak yang bersirkulasi dengan meningkatkan penyerapannya dalam jaringan adiposa putih dan meningkatkan pengeluaran energi, yang kemungkinan berkontribusi pada penyimpanan lemak hati yang lebih rendah [ 101 ]. Sementara mekanisme yang tepat untuk respons termogenik tidak jelas, katekin teh hijau tampaknya awalnya memengaruhi PPARγ, yang berpotensi menyiapkan panggung untuk jaringan lemak yang lebih aktif secara metabolik. Selain itu, ekstrak fenolik dari cranberry dapat meningkatkan aktivitas PRDM16 dalam BAT, tetapi tidak dalam eWAT [ 102 ]. Pengamatan yang dilakukan dengan polifenol apel menunjukkan bahwa peningkatan kapasitas termogenik yang disebabkan oleh asupan adalah hasil dari adaptasi iWAT, bukan dari perubahan BAT atau eWAT [ 103 ]. Disosiasi antara mRNA dan kelimpahan protein ini telah dilaporkan oleh peneliti lain dalam model yang berbeda, yang menunjukkan mekanisme regulasi pasca-transkripsi yang potensial, seperti pergantian protein, mungkin terlibat dalam regulasi UCP1.
Ketergantungan reseptor adrenergik pada pencoklatan jaringan adiposa yang diinduksi flavanol kakao diidentifikasi pada tikus C57BL/6J yang diberi ekstrak kakao secara oral [ 104 ]. Setelah 2 minggu, peningkatan signifikan ekskresi norepinefrin dan adrenalin dan peningkatan ekspresi penanda protein pencoklatan (UCP-1, PGC-1α, PRDM16) diamati. Penghambat β-adrenergik spesifik menunjukkan bahwa peningkatan ekspresi protein ini dapat dicegah setelah pemberian flavanol [ 105 ]. Sementara hubungan antara reseptor adrenergik dan termogenesis yang diinduksi dingin sudah mapan, mekanisme yang mendasari saling ketergantungan mereka dalam pencoklatan jaringan adiposa, memerlukan penjelasan lebih lanjut.
Konsumsi polifenol, baik dalam hal kuantitas dan kualitas, dapat memengaruhi respons tubuh terhadap paparan cahaya. Sebuah studi oleh Navarro-Masip et al. [ 106 ] mengevaluasi bagaimana fotoperiode yang berbeda, yang mensimulasikan perubahan musiman, memengaruhi parameter tubuh dan protein pada tikus Fischer 344 yang diberi makan anggur biasa atau organik. Tikus-tikus tersebut terpapar cahaya selama 6, 12, atau 18 jam per hari. Studi tersebut menemukan bahwa pengeluaran energi menurun pada tikus yang terpapar periode cahaya seperti musim dingin (6 jam), sementara itu meningkat pada tikus yang terpapar periode cahaya yang lebih lama (12 dan 18 jam) [ 106 ]. Selain itu, pencoklatan iWAT hanya diamati pada periode cahaya 12 dan 18 jam, sehingga menimbulkan pertanyaan menarik tentang adaptasi evolusioner untuk termogenesis dan musim.
Ekstrak antosianin dari blueberry dan cranberry mengurangi berat badan dan akumulasi lemak pada tikus yang diberi makanan tinggi lemak, tetapi hanya pada dosis yang lebih tinggi (2%) jus cranberry dapat mengurangi berat badan [ 107 ]. Ekstrak ini dapat bekerja dengan mengurangi penanda inflamasi dan meningkatkan pelepasan SCFA sambil menurunkan produksi LPS melalui modulasi mikrobiota usus, yang berpotensi menawarkan strategi untuk mengelola obesitas dan risiko kesehatannya [ 107 ].
8 Hubungan Evolusioner dan Perspektif Studi Termogenesis dan Polifenol
Meskipun jelas bahwa suhu dingin dapat memicu aktivitas BAT yang ditingkatkan dan pencoklatan WAT, mengapa senyawa fenolik dapat meniru aktivitas ini pada tikus dan manusia cukup menarik. Contoh mencolok pertama yang terlintas dalam pikiran adalah beruang yang berhibernasi. Beruang cokelat menimbun lemak untuk hibernasi dengan melahap buah beri selama berjam-jam, yang mengandung banyak fruktosa dan polifenol, di musim panas dan gugur [ 108 ]. Termogenesis pembakar lemak yang dipromosikan mungkin terkait dengan bagaimana tubuh beruang menangani asam urat, produk sampingan dari metabolisme fruktosa [ 109 ]. Menariknya, baru-baru ini diselidiki bahwa diet tinggi frutosa menekan penyerapan glukosa pada BAT manusia [ 110 ].
Beberapa penelitian mendukung gagasan bahwa senyawa-senyawa ini mengaktifkan jalur transkripsi gen yang mengarah pada peningkatan aktivitas termogenik, termasuk proliferasi mitokondria dan peningkatan metabolisme lipid [ 98 , 103 , 111 ]. Signifikansi evolusioner dari saling ketergantungan ini dan implikasinya bagi hewan-hewan ini saat ini tidak diketahui. Meskipun demikian, model hewan yang tidak berhibernasi, seperti tikus, telah dipelajari secara ekstensif untuk tujuan ini, yang menunjukkan pencoklatan jaringan adiposa sebagai respons terhadap asupan senyawa-senyawa ini. Apakah jalur-jalur ini mewakili sisa-sisa evolusi atau memiliki fungsi adaptif bagi individu-individu ini dan sejauh mana mereka relevan pada manusia juga masih harus dijelaskan. Karena kelimpahan jaringan adiposa coklat (BAT) yang lebih tinggi pada bayi baru lahir manusia dibandingkan dengan orang dewasa, BAT secara luas diakui memainkan peran penting dalam termoregulasi pada bayi, yang tidak dapat melindungi diri dari suhu dingin [ 112 ]. Sebuah penelitian baru-baru ini dengan perenang Skandinavia telah menunjukkan peningkatan aktivitas BAT saat berenang di air dingin [ 113 ].
Di sisi lain, pemberian fruktosa akut, tetapi bukan glukosa, dengan cepat meningkatkan kadar FGF21 pada manusia [ 114 , 115 ]. Gen FGF21 mamalia sangat terkonservasi, memang, hanya ada perbedaan satu atau dua asam amino antara manusia dan gorila atau manusia dan simpanse, masing-masing, dan ada hampir 80% homologi antara FGF21 manusia dan hewan pengerat [ 116 ]. Protein ini, yang banyak diekspresikan di hati, diketahui menginduksi pencoklatan iWAT, melalui induksi PPARα [ 36 ], yang menyebabkan lipolisis. Juga, seperti yang disebutkan, ADBR2 bukan ADBR3 adalah target utama untuk aktivasi farmakologis adiposit coklat manusia. Bagaimana FGF21R berinteraksi dengan reseptor ini untuk menyebarkan sinyal termogenik pada manusia mungkin berbeda dari tikus, yang lebih responsif terhadap ADBR3, secara signifikan.
Sementara artikel-artikel terkini berfokus pada hubungan antara mikrobiota dan FXR sebagai kemungkinan yang bertanggung jawab atas aktivasi termogenik oleh polifenol, peran aktivasi ini dan perbedaan antara metabolit dan struktur ini terkait aktivasi yang lebih efektif belum ditentukan. Sejauh mana mengonsumsi buah beri ini, yang kaya akan senyawa fenolik, dapat menyebabkan pencoklatan jaringan adiposa pada manusia, dan apakah ada korelasi dengan hewan yang mengalami stres dingin, masih harus diklarifikasi.

