
ABSTRAK
Anemia defisiensi besi (IDA) sangat lazim di Afrika sub-Sahara, di mana populasi sering bergantung pada tanaman pokok miskin nutrisi sebagai sumber energi utama mereka. Pisang dataran tinggi Afrika Timur (EAHB), makanan pokok bagi jutaan orang di Uganda dan sebagian besar dataran tinggi Afrika Timur, mengandung zat besi yang tidak mencukupi untuk memenuhi kebutuhan makanan. Berdasarkan konsumsi harian rata-rata dan asupan zat besi yang direkomendasikan, biofortifikasi EAHB dengan lebih dari 18,3 mg/kg DW zat besi dapat secara signifikan mengurangi IDA di komunitas ini. Analisis komprehensif dari 43 genotipe Musa mengungkapkan bahwa, meskipun konsentrasi zat besi dalam bubur buah pisang umumnya rendah, konsentrasi yang jauh lebih tinggi ditemukan dalam bubur spesies liar dan kultivar Fe’i dibandingkan dengan kelompok genom lainnya. Pada jaringan lain, seperti kulit dan biji diploid fertil liar, konsentrasi zat besi secara konsisten lebih tinggi, menggarisbawahi peran penting mineral ini dalam fotosintesis dan perkembangan biji. Analisis genomik dan filogenetik pada lima kultivar pisang terpilih mengidentifikasi 37 gen yang terkait dengan homeostasis zat besi, yang mencakup enam keluarga protein yang berbeda, dan mengungkap perbedaan yang jelas antara Fe’i dan kultivar yang penting secara komersial, Cavendish. Sebuah studi ekspresi gen diferensial khusus jaringan di Cavendish selanjutnya mengidentifikasi regulator utama homeostasis zat besi dalam tanaman ini. Temuan ini menyediakan sumber daya dasar untuk biofortifikasi buah penting ini dan berkontribusi untuk mengatasi tantangan kesehatan global yang terus-menerus.
1 Pendahuluan
Kekurangan zat besi adalah kekurangan nutrisi yang paling umum pada manusia dan penyebab utama anemia, yang menimbulkan risiko signifikan terhadap kesehatan global (Kumar et al. 2022 ). Dengan lebih dari setengah miliar individu diperkirakan menderita IDA dan/atau simpanan zat besi yang menipis, dampak dari kondisi ini tidak dapat diremehkan (Johnson et al. 2011 ). Selain itu, kekurangan zat besi secara tidak proporsional mempengaruhi populasi rentan di wilayah terbatas sumber daya di mana bahkan kondisi ringan dapat mengakibatkan hasil kesehatan yang melemahkan (Finkelstein et al. 2017 ; Detzel dan Wieser 2015 ). Uganda, negara berkembang yang terletak di wilayah Great Lakes di Afrika Timur, menghadapi masalah kesehatan yang substansial karena kekurangan zat gizi mikro dalam makanan, khususnya vitamin A, zat besi, seng, dan yodium. Sekitar 53% anak-anak Uganda di bawah usia lima tahun dan 35% wanita pada usia reproduksi menderita IDA (UBOS 2017 ). Mengatasi tantangan kesehatan masyarakat yang signifikan ini akan sangat penting untuk meningkatkan kesejahteraan dan tumbuh kembang anak, meningkatkan kualitas hidup secara keseluruhan, dan mengurangi angka kematian terkait dalam komunitas ini.
Pisang dataran tinggi Afrika Timur (EAHB) adalah kelompok kultivar Musa yang secara genetik unik , terutama triploid (AAA) tetapi juga mencakup beberapa genotipe diploid (AA), yang dibudidayakan secara luas di wilayah Great Lakes di Afrika Timur (Perrier et al. 2019 ). Pisang ini merupakan makanan pokok di Uganda dan negara-negara tetangga, dengan konsumsi per kapita tertinggi di dunia yang mencapai hingga 300 kg per tahun (Fungo 2010 ). EAHB sangat penting bagi ketahanan pangan regional, menyediakan zat gizi makro dan mikro esensial bagi sekitar 30 juta orang, yang banyak di antaranya rentan terhadap IDA (Perrier et al. 2019 ; Fungo dan Pillay 2011 ). Sayangnya, konsentrasi zat gizi mikro esensial dalam makanan pokok utama ini selalu rendah, berkisar antara 0,7 hingga 6,1 mg/kg berat kering (DW) untuk zat besi dan 0,0 hingga 5,4 mg/kg DW untuk seng (Pillay dan Fungo 2016 ), yang telah memperburuk prevalensi IDA dan kekurangan seng di wilayah tersebut (Matovu 2016 ; Fungo 2010 ).
Biofortifikasi tanaman pangan pokok melalui pemuliaan konvensional atau modifikasi genetik telah terbukti menjadi strategi yang berkelanjutan dan hemat biaya untuk mengatasi kekurangan zat besi. Misalnya, jewawut yang dibiakkan secara konvensional dan diperkaya zat besi (Govindaraj et al. 2019 ), kacang-kacangan (Katsvairo 2014 ), dan beras (Gregorio et al. 2000 ) telah menunjukkan hasil yang menjanjikan dalam mengatasi kekurangan zat besi di India, Rwanda, dan Filipina, masing-masing (Pompano et al. 2022 ; Haas et al. 2016 , 2005 ). Demikian pula, singkong (Narayanan et al. 2019 , 2015 ; Taylor et al. 2016 ), beras (Kawakami et al. 2022 ; Johnson et al. 2011 ), dan gandum (Beasley et al. 2019 ; Connorton et al. 2017 ) telah dimodifikasi secara genetik untuk meningkatkan kandungan zat besi.
Pada tahun 2005, proyek biofortifikasi Banana21, yang didukung oleh Bill & Melinda Gates Foundation, diprakarsai untuk membantu mengatasi kekurangan vitamin A dan zat besi di Uganda melalui peningkatan mikronutrien EAHB menggunakan modifikasi genetik (Paul et al. 2017 ). Pendekatan ini dianggap paling layak karena rendahnya kesuburan pisang yang dapat dimakan, yang membuat pemuliaan konvensional menjadi sulit (Becker et al. 2000 ) dan ketersediaan protokol transformasi untuk banyak kultivar pisang (Khanna et al. 2007 , 2004 ). Namun, tidak seperti tanaman lain seperti padi (Wu et al. 2022 ; Vasconcelos et al. 2017 ), gandum (Connorton dan Balk 2019 ; Bonneau et al. 2016 ) dan barley (Tauris et al. 2009 ), pemahaman saat ini tentang homeostasis mineral pada pisang sangat terbatas (Yadav et al. 2017 ).
Besi tidak disintesis secara biologis di dalam tanaman , tetapi diperoleh dari tanah; karenanya, tanaman telah mengembangkan mekanisme yang rumit untuk mengatur penyerapan dan penyimpanannya untuk mempertahankan homeostasis (Briat et al. 1995 ). Kemajuan yang signifikan telah dibuat dalam meningkatkan biofortifikasi besi di berbagai tanaman melalui rekayasa genetika dengan pemahaman yang mendalam tentang proses molekuler yang terlibat dalam mobilisasi besi dari tanah, penyerapan dari rizosfer, translokasi, dan redistribusi melalui jaringan udara, dan akumulasi dalam bentuk yang tersedia secara hayati di bagian tanaman yang dapat dimakan (Sánchez et al. 2012 ; Morrissey dan Guerinot 2009 ; White dan Broadley 2009 ). Tanaman menggunakan strategi yang kompleks untuk memperoleh besi dari tanah, terutama melalui dua metode: Strategi reduksi I dan Strategi khelasi II (Römheld 1987a ). Monokotil nongraminaceous seperti pisang terutama bergantung pada pengasaman dan reduksi untuk mengakses besi dari tanah menggunakan dua enzim utama. Ini termasuk ferric reductase oxidase (FRO) (Robinson et al. 1999 ), yang mereduksi ion ferric menjadi ion ferrous, dan iron-regulated transporter (IRT) (Vert et al. 2002 ), yang mengangkut besi melintasi sel-sel akar. Enzim-enzim penting lainnya dalam proses ini termasuk nicotianamine synthase (NAS) (Takahashi et al. 2003 ; Shojima et al. 1990 ), yang mengkatalisis produksi nicotianamine untuk pengangkutan besi; yellow stripe-like protein (YSL) yang membantu dalam pergerakan kompleks besi dalam tanaman (Koike et al. 2004 ); vacuolar iron transporter (VIT) yang dikaitkan dengan penyimpanan besi dalam vakuola (Kim et al. 2006 ); dan feritin (Fer) yang memfasilitasi penyimpanan zat besi dalam biji dan daun, membantu mengelola konsentrasi zat besi dan mencegah toksisitas (Theil 2003 ; Theil et al. 1997 ).
Bahasa Indonesia: Pada banyak tanaman pangan hasil rekayasa genetika (GM), dengan Golden Rice yang ditingkatkan β-karoten menjadi contoh yang terkenal, sumber transgen dapat sangat memengaruhi fenotipe yang diinginkan (Ye et al. 2000 ). Demikian pula, buah tanaman pisang Cavendish yang mengekspresikan gen fitoena sintase yang berasal dari pisang Fe’i mengandung konsentrasi ekuivalen β-karoten yang lebih tinggi daripada buah dari tanaman pisang transgenik yang mengekspresikan ortolog jagung secara berlebihan (Paul et al. 2017 ). Oleh karena itu, menyelidiki variasi alami dalam konsentrasi mineral buah di berbagai genotipe Musa , termasuk kultivar liar dan yang dikonsumsi, dapat mengidentifikasi sumber daya genetik berharga yang dapat dimanfaatkan untuk biofortifikasi besi. Studi semacam itu saat ini kurang dalam banyak genotipe pisang. Spesies liar biasanya dianggap sebagai sumber daya genetik yang berharga untuk meningkatkan keragaman genetik tanaman pangan yang dibudidayakan (Brozynska et al. 2016 ). Dengan demikian, menyelidiki spesies M. acuminata Colla (AA) dan M. balbisiana Colla (BB), kerabat liar dari sebagian besar pisang hibrida diploid atau triploid yang dapat dimakan, dapat menginformasikan keputusan masa depan yang dibuat terhadap biofortifikasi pisang. Pisang Fe’i adalah kelompok unik dari kultivar masak M. troglodytarum spp. (T) diploid (TT) dan triploid (TTT) yang ditemukan di wilayah Pasifik, yang mudah dikenali dari getahnya yang merah atau ungu, warna daging kuning-oranye tua, dan sangat terkenal karena kandungan pro-vitamin A yang melimpah (Li et al. 2022 ; Thomson et al. 2022 ; Buah et al. 2016 ). Pisang-pisang ini tidak hanya memiliki signifikansi budaya bagi wilayah Pasifik tetapi juga nilai gizi potensial karena karakteristiknya yang berbeda (Thomson et al. 2022 ). Sampai saat ini, relatif sedikit penelitian yang dilakukan untuk menyelidiki kandungan mineral dalam buah Fe’i (Englberger et al. 2003 ) dan spesies pisang liar, juga belum diukur pada kulit dan biji spesies Musa apa pun .
Pada awal inisiatif Banana21, pemahaman tentang dasar genetik regulasi zat besi di Musa diidentifikasi sebagai langkah penting menuju pengembangan strategi biofortifikasi yang efektif untuk tanaman pangan pokok ini. Studi ini bertujuan untuk menetapkan apakah pisang memiliki potensi genetik untuk mengakumulasi dan mengatur konsentrasi zat besi yang signifikan secara gizi dalam buahnya. Untuk memandu upaya ini, ambang batas konsentrasi zat besi buah minimum—didefinisikan sebagai “target”—yang diperlukan untuk berdampak signifikan pada populasi yang bergantung pada pisang sebagai makanan pokok telah dihitung. Untuk mengidentifikasi sumber genetik potensial untuk biofortifikasi, konsentrasi zat besi dan mineral lainnya dalam buah dari 33 genotipe Musa diukur, dan variasi alaminya ditentukan dan dibandingkan dengan 10 kultivar yang umum ditanam di Uganda. Lebih jauh, gen kunci yang terlibat dalam homeostasis zat besi diidentifikasi dalam kultivar yang relevan, dan profil evolusi dan ekspresinya dianalisis, memberikan wawasan tentang kontribusi potensial mereka terhadap peningkatan kandungan zat besi.
2 Bahan dan Metode
2.1 Pengumpulan dan Penanganan Sampel Buah Pisang
Berbagai macam 33 kultivar Musa dipilih yang tumbuh dalam koleksi plasma nutfah yang dikelola oleh Queensland Department of Primary Industries (QDPI), South Johnstone Research Station (SJRS, Australia). Buah hijau penuh (FG) yang matang dipanen dari bagian tengah setiap tandan setidaknya tiga kali selama periode 4 tahun dari tanaman sehat yang berumur sama yang dibudidayakan dalam kondisi lapangan yang sama. Dalam kasus variabilitas yang tinggi antara ulangan, pengambilan sampel ditingkatkan hingga enam ulangan. Untuk tiga kultivar Fe’i, Asupina, Karat, dan Taweyawa, sampel hanya dapat dikumpulkan satu kali karena pengelompokan genotipe ini tidak konsisten. Buah diterima di laboratorium di Brisbane dalam waktu 48 jam setelah panen, difoto, diukur, dan diproses segera. Bobot tandan dikumpulkan dari berbagai sumber literatur dan dirata-ratakan (Smith et al. 2018 ; Daniells dan Hamill 2004 ; Daniells dan Bryde 2001 ; Arnaud dan Horry 1997 ; Daniells 1986 ). Selain itu, buah FG juga dipanen seperti yang dijelaskan sebelumnya dari 10 kultivar yang ditanam di 13 distrik di Uganda, yang mewakili semua zona penanaman pisang agroekologi di negara tersebut.
2.2 Pengolahan Sampel Buah Pisang untuk Analisis dan Penyimpanan Mineral
Buah dicuci dengan air Milli-Q, dikeringkan, dipisahkan menjadi kulit, daging buah, dan biji (bila tersedia), dipotong dadu menggunakan pisau keramik, ditempatkan dalam tabung polipropilena (PP) 50 mL, dan disimpan pada suhu -80°C sebelum dikeringkan dalam suhu beku. Selanjutnya, sampel daging buah digiling menjadi bubuk halus menggunakan lumpang dan alu, diayak menggunakan kasa plastik 0,2 mm, dan disimpan dalam tabung PP 50 mL tertutup dengan tutup ulir polietilena berdensitas tinggi (HDPE) pada suhu ruangan (21°C–23°C). Sampel kulit dan biji digiling menggunakan Mixer Mill MM 400 (Retsch) dan tabung penggiling karbida tungsten dan disimpan seperti yang dijelaskan sebelumnya. Bila diperlukan, sampel pisang dimatangkan secara artifisial pada suhu 20°C dalam wadah polietilena buram tertutup rapat berkapasitas 50 L menggunakan ~10 L etilena 4,5% dalam CO 2 (RIPEGAS, BOC) hingga Tahap 6 yang matang sepenuhnya tercapai seperti ditunjukkan dalam Gambar S1 (USDA, AMS, FVP, dan SCI 2001 ).
2.3 Pencernaan Sampel dan Analisis Konsentrasi Mineral Mikronutrien
Untuk pengukuran konsentrasi mineral, 200 mg sampel kering dicerna dengan asam pada dasarnya seperti yang dijelaskan oleh Wheal et al. ( 2011 ). Setiap sampel diproses dalam rangkap tiga untuk analisis mineral. Konsentrasi mineral diukur menggunakan spektrometri emisi optik plasma yang digabungkan secara induktif (ICP-OES) pada Perkin Elmer ICP-OES 8300DV, yang dikonfigurasi dan protokol diterapkan seperti yang dijelaskan oleh Pepper et al. ( 2021 ). Kekokohan, akurasi, dan presisi metode diverifikasi dengan analisis berkelanjutan dari sampel pisang tipe liar referensi bersertifikat (CRM-IPE 929) yang awalnya dianalisis di Wageningen Evaluating Programs for Analytical Laboratories (WEPAL). Sampel dianalisis menggunakan waktu integrasi 0,15 detik dengan tiga replikasi. Konsentrasi dinyatakan dalam mg/kg berdasarkan berat kering (DW). Sampel diperiksa untuk mengetahui konsentrasi tinggi logam berat dan kontaminan dan diuji ulang jika diduga ada kontaminasi.
2.4 Spektroskopi Fluoresensi Sinar-X Mikro (μ-XRF)
Potongan melintang tipis buah segar dari Zebrina disiapkan menggunakan pisau keramik. Pemetaan unsur μ-XRF dilakukan di Centre for Microscopy and Microanalysis (University of Queensland) dengan ATLAS X micro XRF Spectrometer (IXRF Systems, AS). Pengukuran dilakukan pada suhu ruangan (~20°C) menggunakan sumber sinar-X anoda Molibdenum 25 μm pada 40–50 kV, 1000 μA, dengan waktu naik 0,25 μs dan waktu tinggal per piksel 50–100 ms. Silicon Drift Detector (SDD) digunakan untuk menangkap sinyal fluoresensi penuh. Pemetaan unsur dibuat menggunakan paket perangkat lunak IXRF Iridium Ultra dan GeoPIXE, menggunakan jumlah hitungan pada posisi puncak utama untuk setiap unsur. Masing-masing gambar diekspor ke ImageJ sebagai file TIFF skala abu-abu 8-bit, dinormalisasi secara internal sehingga setiap gambar mencakup rentang dinamis penuh, dan gambar akhir diwarnai menggunakan Imaris 9.3 (instrumen Oxford).
2.5 Analisis Statistik
Semua perbandingan statistik dibuat menggunakan R (ver. 3.5.0, R Foundation for Statistical Computing, Vienna, Austria). Normalitas dan homogenitas varians diuji dengan uji D’agostino-Pearson dan Levene, masing-masing. Untuk studi retensi, uji Mann–Whitney U digunakan untuk menguji varians kedua kelompok. Dalam semua contoh lainnya, analisis varians dilakukan dengan menggunakan uji Kruskal–Wallis H non-parametrik, dengan beberapa perbandingan dibuat dengan uji post hoc Dunn dan nilai p disesuaikan dengan metode Benjamini–Hochberg. Signifikansi statistik dinyatakan pada tingkat kepercayaan 95% ( p > 0,05) untuk semua pengujian. Pengujian ini dilakukan dengan basis R dan paket fBasics ver. 3042.89.2, DescTools ver. 0.99.45, FSA ver. 0.9.3, dan car ver. 3.1-0. Grafik korelasi dan korelogram Pearson dibuat menggunakan corrplot ver. 0.92. Statistik estimasi dan plot Gardner–Altman dibuat menggunakan dabestr ver. 0.2.5. Analisis komponen utama (PCA) dan biplot serta pemodelan selanjutnya dibuat menggunakan factoextra ver. 1.0.6.
2.6 Identifikasi dan Penamaan Gen Homeostasis Besi Dari DH Pahang
Bahasa Indonesia : Untuk mengidentifikasi gen yang terkait dengan homeostasis besi ( Fer , FRO , IRT , NAS , VIT , dan YSL ), urutan asam amino dari gen-gen yang berkarakterisasi baik ini di Arabidopsis thaliana dan Oryza sativa (Tabel S1 ) digunakan sebagai kueri untuk pencarian BLAST terhadap genom pisang DH Pahang ver. 4 yang dianotasi yang tersedia di Banana Genome Hub (Belser et al. 2021 ). Ketika urutan pisang yang cocok secara signifikan lebih panjang daripada padanannya di A. thaliana dan O. sativa , gen putatif dari daerah kromosom yang sama dalam rakitan DH Pahang ver. 2 tetapi dengan ukuran urutan yang benar disubstitusikan (Martin et al. 2016 ). Setiap gen putatif homeostasis besi Musa yang teridentifikasi diberi nama unik menggunakan deskripsi rakitan sebagai default. Urutan homolog dengan deskripsi perakitan generik dibedakan dengan perluasan yang terdiri dari (1) angka yang dimulai dengan angka serendah mungkin dan dalam urutan lokasi kromosom dan (2) huruf (a, b, c…) untuk setiap paralog dalam urutan lokasi pada kromosom yang sama.
2.7 Prediksi Gen Homeostasis Besi pada Kultivar Lain
Bahasa Indonesia: Menggunakan Liftoff ver. 1.6.3 (Shumate dan Salzberg 2021 ), urutan gen homeostasis besi putatif yang diidentifikasi dalam rakitan DH Pahang ver. 4 dan ver. 2 (Tabel S1 ) digunakan untuk memprediksi urutan homolog dan ortolog dari empat rakitan kultivar Musa dan dua Ensete (outgroup). Awalnya, urutan putatif yang diidentifikasi dalam anotasi ver. 2 menggantikan rekan rakitan ver. 4 yang ukurannya salah menggunakan Liftoff. Selanjutnya, semua urutan diekstraksi dari anotasi hibrida ini. Rakitan genom dari kultivar yang dipilih diunduh dari Banana Genome Hub (Droc et al. 2013 ), dan urutan gen ortolog dianotasi menggunakan anotasi hibrida yang baru dibuat. Semua kultivar/aksesi/spesies yang dipilih untuk analisis dan singkatan selanjutnya yang digunakan untuk merujuknya dapat ditemukan di Tabel S2 .
2.8 Penyelarasan Urutan dan Analisis Filogenetik
Untuk setiap keluarga gen pengatur zat besi ( Fer , FRO , IRT , NAS , VIT , dan YSL ), semua sekuens homolog dan ortolog yang diidentifikasi diterjemahkan, diperiksa untuk redundansi, dan disesuaikan secara manual. Ini mengoreksi kesalahan Liftoff, seperti sekuens asing di hulu dari situs awal metionina sejati untuk memastikan penyelarasan dengan kerangka baca terbuka (ORF) dari sekuens anotasi hibrida. Untuk analisis filogenetik, sekuens diselaraskan menggunakan ClustalW ver. 2.1 (Thompson et al. 1994 ) untuk menghasilkan pohon tingkat keluarga protein atau pohon tingkat protein individual. Untuk pohon tingkat keluarga protein, semua sekuens homolog dalam setiap keluarga protein pertama-tama dirangkai untuk setiap kultivar pisang sebelum penyelarasan. Urutan dari Fer2, Fer3b, FRO2b, VIT1b, YSL6a, YSL8, YSL12e, dan YSL12g dikeluarkan dari penggabungan karena kodon stop prematur atau multipel (selanjutnya disebut sebagai “non-fungsional”). Selain itu, karena VIT1a dan VIT4 tidak ada dalam perakitan Utafan, urutan ortolog yang sesuai dari semua kultivar lain dihilangkan sebelum penggabungan. Mengingat jumlah terbatas urutan IRT yang diidentifikasi, semuanya dipertahankan, termasuk yang tidak berfungsi. Protein dari perakitan Ensete ventricosum “Eg” dan E. glaucum “Ev” digabungkan untuk digunakan sebagai “outgroup.” Analisis filogenetik dari urutan protein individu dan yang digabungkan dilakukan dengan menggunakan kemungkinan maksimum dan Kriteria Informasi Akaike sebagaimana diterapkan dalam ModelFinder dalam IQ-TREE ver. 2.2.6 (Minh et al. 2020 ; Kalyaanamoorthy et al. 2017 ). Pohon konsensus akhir divisualisasikan dengan FigTree ver. 1.4.4.
2.9 Analisis RNA-Seq
Data RNA-seq internal dari dua replikasi buah biologis Cavendish (masing-masing satu dari kultivar Grande Naine dan Williams) dan tiga replikasi biologis sampel daun dan akar kultivar Grande Naine digunakan untuk analisis ekspresi gen diferensial. Kualitas pustaka RNA-seq dinilai menggunakan Fastqc ver. 0.11.2 (Andrews 2010 ) sebelum penyelarasan. Menggunakan komputer berkinerja tinggi (HPC), sekuens diselaraskan dengan anotasi hibrida yang dijelaskan sebelumnya menggunakan kallisto ver. 0.46.1 (Bray et al. 2016 ) dan perkiraan jumlah bacaan dihitung. Jumlah bacaan dinormalisasi antara sampel dengan DeSeq2 ver. 1.41.0, data replikasi dirata-ratakan, dan outlier dihapus. Pola ekspresi relatif direpresentasikan pada peta panas yang dibangun dengan paket R pheatmap ver. 1.0.12.
3 Hasil dan Pembahasan
3.1 Target Biofortifikasi untuk Dampak Gizi yang Signifikan
Target konsentrasi zat besi pada buah pisang dihitung untuk menyediakan setidaknya 50% dari estimasi kebutuhan rata-rata (EAR) (Bouis et al. 2011 ; White dan Broadley 2009 ). Kebutuhan harian zat besi yang diserap bervariasi tergantung pada usia dan jenis kelamin populasi target (Tabel 1 ); meskipun demikian, penyerapan dipengaruhi oleh tiga faktor independen: asupan makanan (g/hari), retensi (kehilangan melalui pemrosesan), dan bioavailabilitas (Bouis et al. 2011 ; White dan Broadley 2009 ).
| Kelompok umur (tahun) | EAR untuk zat besi (mg/hari) a | Kebutuhan zat besi yang diserap (μg/hari) b | Zat besi yang diserap diperlukan untuk memenuhi 50% EAR (μg/hari) | Konsumsi pisang (jari/hari) | Konsumsi pisang (g FW/hari) c | Konsumsi pisang (gr DW/hari) d | SM 50 (mg/kg berat badan) |
|---|---|---|---|---|---|---|---|
| Bayi | |||||||
| 7–12 bulan | 7 | tahun 1260 | 630 | 1 | 120 | 30 | 61.8 |
| Anak-anak | |||||||
| 1–3 | 4 | 560 | 280 | 1.5 | 180 | 45 | 18.3 |
| 4–8 | 4 | 720 | 360 | 2.0 | 240 | 60 | 17.6 |
| 9–13 | 6 | tahun 1080 | 540 | 3.0 | 360 | 90 | 17.6 |
| 14–18 | 8 | tahun 1440 | 720 | 4.0 | 480 | 120 | 17.6 |
| Pria | |||||||
| 19+ | 6 | tahun 1080 | 540 | 6.0 | 720 | 180 | 8.8 |
| Wanita | |||||||
| 19–50 | 8 | tahun 1440 | 720 | 5.0 | 600 | 150 | 14.1 |
| 51+ | 5 | 900 | 450 | 5.0 | 600 | 150 | 8.8 |
| Kehamilan | |||||||
| 14–18 | 23 | 4140 | tahun 2070 | 4.0 | 480 | 120 | 50.7 |
| 19–50 | 22 | 3960 | tahun 1980 | 5.0 | 600 | 150 | 38.8 |
| Laktasi | |||||||
| 14–18 | 7 | tahun 1260 | 630 | 4.0 | 480 | 120 | 15.4 |
| 19–50 | 6.5 | tahun 1170 | 585 | 5.0 | 600 | 150 | 11.5 |
Singkatan: DW, berat kering; EAR, perkiraan kebutuhan rata-rata; FW, berat segar. Catatan: bc 50 , konsentrasi zat besi buah pisang yang dibutuhkan untuk mencapai 50% dari kebutuhan zat besi, dinyatakan dalam mg/kg DW dan dihitung sebagai berikut: bc 50 = , I = zat besi yang diserap yang dibutuhkan untuk memenuhi 50% EAR (μg/hari); C = konsumsi pisang (g DW/hari); R = retensi pemasakan sebesar 100% (Tabel S3 ); A = penyerapan sebesar 34% (García et al. 2015 ). EAR ditetapkan dengan memodelkan komponen kebutuhan zat besi, memperkirakan kebutuhan zat besi yang diserap pada persentil ke-50 dengan menggunakan batas atas penyerapan zat besi sebesar 14% untuk kelompok anak usia 1–3 tahun dan 18% untuk kelompok usia lainnya, dengan pembulatan (NHMRC et al. 2006 ). b Berdasarkan 14% dan 18% bioavailabilitas dalam makanan anak usia 1–3 tahun dan kelompok usia lainnya. c Berdasarkan berat rata-rata jari pisang 120 g FW. d Dengan asumsi 25% bahan kering.
Konsumsi pisang masak di Uganda adalah yang tertinggi di dunia, dengan laporan melebihi 1 kg per orang per hari (Fungo 2010 ). Dengan tidak adanya referensi konsumsi pisang yang andal dan terperinci untuk kelompok usia yang berbeda di Uganda, konsumsi pisang yang dimasak (hidangan nasional, matoke) diperkirakan secara konservatif dan disesuaikan untuk setiap kelompok usia (Tabel 1 ) (García et al. 2015 ; NHMRC et al. 2006 ). Di Uganda, matoke disiapkan dengan merebus atau mengukus buah pisang yang sudah dikupas setidaknya selama satu jam sebelum dihaluskan. Ketika kandungan zat besi dan seng dari bubur buah pisang Cavendish yang direbus selama 1 jam dalam air dibandingkan dengan yang setara dengan yang tidak direbus, tidak ada kerugian signifikan ( p > 0,05) yang dapat dikaitkan dengan teknik pemrosesan ini (Tabel S3 ). Akhirnya, pekerjaan sebelumnya yang bekerja sama dengan laboratorium kami telah menunjukkan tingkat penyerapan sebesar 34% dari pisang Cavendish yang dimasak (García et al. 2015 ). Dari informasi ini, diperkirakan bahwa antara 8,8 hingga 18,3 mg/kg DW zat besi diperlukan dalam pisang biofortifikasi untuk mencapai 50% dari EAR zat besi dalam berbagai kelompok umur (Tabel 1 ). Mempertimbangkan bahwa kandungan zat besi dalam buah kultivar EAHB secara konsisten dilaporkan rata-rata sekitar 10 mg/kg DW (Matovu 2016 ), kelompok umur seperti pria dan wanita dewasa yang berusia di atas 50-an mungkin sudah mendapat manfaat besar dari pisang ini karena jumlah konsumsi pisang harian yang besar atau berkurangnya kebutuhan zat besi yang diserap. Di sisi lain, anak-anak dan wanita usia subur, dengan kebutuhan zat besi yang lebih tinggi (kg untuk kg) dan/atau kebiasaan konsumsi yang jauh lebih kecil, jelas tidak mendapat manfaat dari kultivar saat ini. Perhitungan kami ditujukan untuk mendukung kedua kelompok ini yang khususnya berisiko kekurangan zat gizi mikro karena kebutuhan nutrisi mereka yang lebih tinggi untuk pertumbuhan dan kesehatan reproduksi (Bouis et al. 2011 ). Populasi anak-anak (1–3 tahun) sejauh ini merupakan kelompok yang paling rentan terhadap segala jenis kekurangan gizi dan khususnya terhadap IDA. Menurut asumsi dan perhitungan yang dibuat, mengonsumsi pisang dengan konsentrasi zat besi minimal 18,3 mg/kg DW dapat menyediakan sekitar 50% dari EAR zat besi untuk kelompok usia ini (Tabel 1) .), yang seharusnya menghasilkan peningkatan signifikan dalam kesehatan dan perkembangan secara keseluruhan. Demikian pula, kelompok paling rentan berikutnya, wanita usia reproduksi, juga dapat memperoleh 50% dari EAHB zat besi dengan mengonsumsi pisang dengan konsentrasi zat besi minimal 14,1 mg/kg DW. Secara keseluruhan, data menunjukkan bahwa agar sebagian besar kelompok usia dapat memperoleh manfaat dari EAHB biofortifikasi zat besi, konsentrasi minimum yang dibutuhkan akan berada dalam kisaran 14,1 hingga 18,3 mg/kg DW, dengan ambang batas yang lebih tinggi mewakili tujuan yang lebih pragmatis dan bermanfaat, yang bertujuan untuk membantu anggota masyarakat yang paling rentan terhadap gizi.
3.2 Konsentrasi Zat Besi dan Seng dalam Jaringan Buah Kultivar Musa
Sebagian besar penelitian tentang konsentrasi mineral buah pada pisang difokuskan pada kultivar yang umum dikonsumsi, dengan konsentrasi zat besi yang dilaporkan jarang melebihi 10 mg/kg. Sampai saat ini, tidak ada kultivar pisang yang dilaporkan dengan konsentrasi zat besi buah yang mendekati target terhitung yang diperlukan untuk biofortifikasi. Untuk menilai konsentrasi zat besi buah pada kultivar pisang yang lebih beragam dan kurang dikenal, konsentrasi zat besi dan seng diukur dalam buah dari 33 kultivar Musa yang ditanam dalam satu koleksi plasma nutfah di Australia. Kultivar ini berkisar dari pisang pencuci mulut yang banyak dikonsumsi seperti Cavendish (AAA) hingga kultivar yang lebih eksotis, termasuk pisang masak, pisang raja, pisang Fe’i, dan diploid fertil liar, yang mencakup berbagai macam kelompok ploidi dan genom (Tabel 2 ). Keragaman fenotipe buah ditunjukkan pada Gambar 1 , dengan diploid fertil liar menghasilkan buah yang jauh lebih kecil berisi biji keras, berbeda dengan kultivar lainnya. Untuk melengkapi penelitian ini, satu set besar sampel bubur buah juga dikumpulkan dari pisang yang ditanam di pertanian di Uganda. Ini termasuk empat kultivar EAHB sejati dan satu hibrida EAHB serta satu pisang raja dan empat varietas makanan penutup yang disertakan untuk tujuan perbandingan dengan tanaman yang ditanam di Australia (Tabel 2 ).
| Kelompok genom | Subgrup | Kultivar/varietas | N | Asal | Bubur | Mengupas | Benih | Lebar jari (cm) | Panjang jari (cm) | Berat tandan b (kg) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Besi (mg/kg) | Seng (mg/kg) | Besi (mg/kg) | Seng (mg/kg) | Besi (mg/kg) | Seng (mg/kg) | ||||||||
| AA (Diploid yang dapat dimakan) | Tidak tersedia | Paka | 3 | Bahasa Indonesia: Australia | 8,4 ± 0,4 | 8,0 ± 1,0 | 22,1 ± 2,2 | 23,4 ± 1,1 | — | — | 3,2 ± 0,1 | 21,1 ± 0,3 | 16,3 ± 3,3 hari |
| Lebih manis | Terema | 3 | Bahasa Indonesia: Australia | 8,2 ± 1,4 | 8,2 ± 1,3 | 35,4 ± 7,7 | 51,5 ± 6,4 | — | — | 2,9 ± 0,2 | 9,9 ± 0,4 | 9,3 ± 5,2 c,g , c,g | |
| Bahasa Inggris | Bahasa Cavendish | Naine Besar | 3 | Bahasa Indonesia: Australia | 9,1 ± 1,5 | 7,2 ± 0,6 | 34,9 ± 9,8 | 34,2 ± 2,7 | — | — | 3,6 ± 0,1 | 18,6 ± 2,8 | 37,5 ± 7,1 hari |
| Bahasa Cavendish | Williams | 6 | Bahasa Indonesia: Australia | 9,1 ± 1,0 | 6,4 ± 1,3 | 24,4 ± 7,3 | 30,5 ± 7,6 | — | — | 3,6 ± 0,1 | 18,4 ± 2,9 | 37,5 ± 7,1 f,g , f,g | |
| Bahasa Cavendish | Kurcaci Cavendish | 1 | Bahasa Indonesia: Australia | 11,9 ± 0,7 | 6,2 ± 0,5 | 35,6 ± 0,3 | 70,7 ± 1,7 | — | — | 3,6 ± 0,0 | 17,4 ± 0,0 | 37,5 ± 17,7 derajat | |
| Bahasa Cavendish | D5 | 2 | Bahasa Indonesia: Australia | 11,8 ± 0,0 | 8,0 ± 1,3 | 27,2 ± 1,8 | 33,4 ± 1,1 | — | — | 3,6 ± 0,1 | 20,8 ± 0,6 | 26,3 ± 5,3 jam | |
| Bahasa Cavendish | DPM25 | 1 | Bahasa Indonesia: Australia | 8,6 ± 0,4 | 6,3 ± 0,1 | 30,0 ± 0,7 | 39,2 ± 1,7 | — | — | 3,5 ± 0,0 | 21,8 ± 0,0 | — | |
| Bahasa Cavendish | Valery | 1 | Bahasa Indonesia: Australia | 9,4 ± 0,2 | 9,4 ± 0,7 | 39,9 ± 0,3 | 54,1 ± 0,7 | — | — | 3,1 ± 0,0 | 24,9 ± 0,0 | 25,4 ± 3,7 hari | |
| Bahasa Cavendish | Kurcaci Cavendish | 27 | Uganda | 10,9 ± 1,2 | 6,7 ± 0,9 | — | — | — | — | — | — | — | |
| Besar Michel | Bogoya | 30 | Uganda | 11,1 ± 1,2 | 5,7 ± 0,5 | — | — | — | — | — | — | — | |
| Sungai | Kluai Hom Thong Mokho | 4 | Bahasa Indonesia: Australia | 8,8 ± 1,2 | 6,6 ± 1,2 | 23,2 ± 7,1 | 36,1 ± 3,9 | — | — | 3,3 ± 0,3 | 17,1 ± 1,6 | 18,4 ± 8,0 d,f , d,f | |
| Lakatan | Lakatan | 4 | Bahasa Indonesia: Australia | 9,3 ± 1,8 | 7,6 ± 1,4 | 25,9 ± 8,1 | 22,6 ± 2,5 | — | — | 3,4 ± 0,1 | 17,7 ± 2,5 | 23,1 ± 2,1 hari | |
| Tidak tersedia | Pisang Susu | 4 | Bahasa Indonesia: Australia | 9,2 ± 0,6 | 6,2 ± 0,3 | 19,6 ± 3,1 | 32,7 ± 1,8 | — | — | 3,8 ± 0,2 | 13,2 ± 1,3 | 14,7 ± 6,0 hari | |
| Merah | Dacca Merah | 3 | Bahasa Indonesia: Australia | 7,9 ± 0,9 | 6,4 ± 1,7 | 20,9 ± 3,7 | 35,2 ± 6,2 | — | — | 4,3 ± 0,3 | 15,5 ± 0,9 | 23,5 ± 5,6 detik | |
| Bahasa Ibota | Yangambi km5 | 3 | Bahasa Indonesia: Australia | 8,6 ± 1,7 | 6,2 ± 0,9 | 24,9 ± 4,7 | 49,0 ± 7,7 | — | — | 3,0 ± 0,3 | 12,8 ± 3,3 | 23,8 ± 10,5 hari | |
| AAA-EAHB | Mutika/Lujugira | Kisanak | 24 | Uganda | 11,4 ± 1,0 | 8,2 ± 1,5 | — | — | — | — | — | — | — |
| Hibrida Mutika/Lujugira | M9 | 24 | Uganda | 11,6 ± 1,4 | 7,5 ± 2,9 | — | — | — | — | — | — | — | |
| Mutika/Lujugira | Bahasa Inggris Mbwazirume | 39 | Uganda | 10,7 ± 2,8 | 7,9 ± 1,6 | — | — | — | — | — | — | — | |
| Mutika/Lujugira | Nakitembe | 33 | Uganda | 10,1 ± 2,3 | 8,0 ± 1,5 | — | — | — | — | — | — | — | |
| Mutika/Lujugira | Bahasa Indonesia: Mpologoma | 33 | Uganda | 10,5 ± 2,2 | 7,1 ± 0,7 | — | — | — | — | — | — | — | |
| Bahasa Inggris AAB | buah pome | Jari Wanita | 4 | Bahasa Indonesia: Australia | 8,1 ± 0,5 | 4,8 ± 1,2 | 44,6 ± 15,7 | 43,0 ± 2,1 | — | — | 3,4 ± 0,3 | 13,0 ± 0,4 | 20,0 ± 14,1 detik |
| Sukali Ndiizi | Kamaramasenge | 27 | Uganda | 9,4 ± 1,6 | 5,5 ± 1,0 | — | — | — | — | — | — | — | |
| Pisang raja | Pisang raja Perancis kerdil | 4 | Bahasa Indonesia: Australia | 8,8 ± 0,5 | 5,2 ± 0,7 | 27,8 ± 4,3 | 38,1 ± 14,9 | — | — | 3,8 ± 0,1 | 21,3 ± 0,5 | 21,2 ± 2,9 hari | |
| Pisang raja | Gonja Nakatasese | 28 | Uganda | 12,1 ± 2,6 | 6,0 ± 1,7 | — | — | — | — | — | — | — | |
| Laknau | Mangat | 3 | Bahasa Indonesia: Australia | 8,1 ± 2,2 | 4,9 ± 1,3 | 22,1 ± 4,4 | 39,6 ± 3,0 | — | — | 4,0 ± 0,2 | 23,2 ± 0,6 | 13,8 ± 4,4 misalnya , misalnya | |
| Bahasa Maoli/Popoulu | Pisang raja Pasifik | 3 | Bahasa Indonesia: Australia | 6,9 ± 0,4 | 7,1 ± 0,6 | 20,1 ± 1,9 | 29,1 ± 8,5 | — | — | 4,7 ± 0,4 | 19,9 ± 1,3 | 18,5 ± 2,1 hari | |
| Pisang Raja | Pisang Raja | 3 | Bahasa Indonesia: Australia | 7,6 ± 1,7 | 5,9 ± 0,8 | 28,2 ± 5,8 | 40,1 ± 10,2 | — | — | 3,5 ± 0,2 | 15,6 ± 1,0 | 19,8 ± 5,0 derajat | |
| Sutra | Gula | 3 | Bahasa Indonesia: Australia | 6,3 ± 1,1 | 4,6 ± 0,5 | 22,5 ± 10,9 | 31,9 ± 6,6 | — | — | 3,3 ± 0,2 | 12,0 ± 1,6 | 16,0 ± 7,9 detik | |
| Iholena | Tigua | 4 | Bahasa Indonesia: Australia | 7,6 ± 0,3 | 9,5 ± 1,3 | 30,1 ± 20,4 | 44,5 ± 11,7 | — | — | 4,1 ± 0,5 | 17,2 ± 2,6 | 17,9 ± 5,7 gram | |
| ABB | Tidak tersedia | Da Jiao | 4 | Bahasa Indonesia: Australia | 8,0 ± 1,3 | 8,0 ± 1,1 | 45,3 ± 23,4 | 31,3 ± 9,9 | — | — | 3,9 ± 0,2 | 16,5 ± 1,7 | 17,4 ± 2,0 hari |
| Pisang Awak | Ducasse | 4 | Bahasa Indonesia: Australia | 8,4 ± 0,9 | 5,1 ± 0,7 | 45,3 ± 7,1 | 52,1 ± 4,7 | — | — | 3,7 ± 0,1 | 14,8 ± 1,1 | 28,3 ± 5,8 detik | |
| bulan | bulan | 3 | Bahasa Indonesia: Australia | 8,4 ± 1,0 | 6,8 ± 0,9 | 26,7 ± 13,2 | 24,0 ± 8,9 | — | — | 4,3 ± 0,5 | 17,0 ± 2,6 | 15,1 ± 17,8 hari | |
| Saba | Pisang Gajih Merah | 3 | Bahasa Indonesia: Australia | 8,8 ± 2,0 | 8,4 ± 1,9 | 35,4 ± 3,2 | 28,0 ± 12,3 | — | — | 3,9 ± 0,4 | 15,4 ± 0,3 | 22,9 ± 2,6 hari | |
| Bluggoe | Bahasa Kivu | 21 | Uganda | 11,2 ± 1,7 | 6,9 ± 0,8 | — | — | — | — | — | — | — | |
| Fei (T) | Tidak tersedia | Asupina (TTT) | 1 | Bahasa Indonesia: Australia | 13,3 ± 0,4 | 10,7 ± 0,5 | 21,1 ± 0,5 | 19,6 ± 0,1 | — | — | 4,4 ± 0,0 | 16,8 ± 0,0 | 10,2 ± 5,1 gram, jam , gram, jam |
| Tidak tersedia | Karat (TTT) | 1 | Bahasa Indonesia: Australia | 14,4 ± 0,3 | 24,8 ± 1,6 | 18,0 ± 0,4 | 37,8 ± 1,2 | — | — | 4,0 ± 0,0 | 12,2 ± 0,0 | 12,5 ± 3,5 tahun | |
| Tidak tersedia | Taweyawa (TT) | 1 | Bahasa Indonesia: Australia | 15,7 ± 0,2 | 24,3 ± 0,5 | 19,8 ± 0,4 | 32,4 ± 0,1 | — | — | 4,9 ± 0,0 | 23,5 ± 0,0 | 3,0 ± 1,4 tahun | |
| Tidak tersedia | Utafan (TT) | 3 | Bahasa Indonesia: Australia | 11,6 ± 1,5 | 12,8 ± 3,0 | 24,9 ± 9,7 | 27,3 ± 10,2 | — | — | 6,0 ± 0,8 | 27,2 ± 2,8 | 22,2 ± 2,6 gram,jam , gram,jam | |
| Tidak tersedia | Ya (AT) sebuah | 3 | Bahasa Indonesia: Australia | 10,6 ± 0,5 | 7,0 ± 1,2 | 35,6 ± 4,0 | 65,5 ± 3,8 | — | — | 3,0 ± 0,6 | 15,4 ± 4,6 | 15,7 ± 4,0 tahun | |
| Liar (Diploid Fertil) | M.a.ssp.malaccensis ( AA ) | 845 | 6 | Bahasa Indonesia: Australia | 12,5 ± 0,8 | 9,1 ± 1,0 | 34,3 ± 5,4 | 21,8 ± 6,0 | 37,4 ± 4,8 | 40,6 ± 11,8 | 2,0 ± 0,2 | 8,5 ± 1,0 | 3,0 ± 0,9 jam |
| M.a.ssp.microcarpa ( AA ) | Kalimantan | 4 | Bahasa Indonesia: Australia | 16,1 ± 2,6 | 31,6 ± 16,5 | 36,4 ± 5,0 | 59,7 ± 9,2 | 38,0 ± 12,9 | 60,2 ± 12,8 | 1,7 ± 0,1 | 9,5 ± 1,1 | 2,5 ± 1,4 hari | |
| M.a.ssp.zebrina ( AA ) | Zebrina | 3 | Bahasa Indonesia: Australia | 28,9 ± 7,8 | 27,0 ± 8,3 | 31,5 ± 8,3 | 60,2 ± 21,9 | 61,3 ± 16,1 | 65,8 ± 14,0 | 2,1 ± 0,1 | 8,2 ± 0,9 | 2,4 ± 1,2 jam | |
| M.b. (BB) | Lal Velchi | 4 | Bahasa Indonesia: Australia | 15,5 ± 6,0 | 23,0 ± 14,4 | 18,0 ± 6,3 | 20,3 ± 5,0 | 21,4 ± 7,1 | 53,7 ± 33,5 | 3,2 ± 0,2 | 12,9 ± 0,4 | 8,9 ± 3,9 jam | |
Catatan: Data dinyatakan sebagai rata-rata n ± simpangan baku (SD). Bila n = 1, SD dihitung dari replikasi teknis. Data Fe dan Zn dihitung berdasarkan berat kering. Singkatan: Fe, besi; M. a ., Musa acuminata ; M. b ., Musa balbisiana ; n , jumlah replikasi biologis; N/A, tidak berlaku; Zn, seng. hibrida M. acuminata (A) × M. troglodytarum (T). b Berat tandan dihimpun dari literatur dan dirata-ratakan. dan Daniells ( 1986 ). d Daniells dan Bryde ( 2001 ). e Daniells, JW komunikasi pribadi. f Smith dan kawan-kawan ( 2018 ). g Arnaud dan Horry ( 1997 ). h Daniells dan Hamill ( 2004 ).

Awalnya, konsentrasi zat besi dan seng dalam buah hijau penuh (FG) dianalisis untuk subset 23 kultivar yang mewakili semua enam kelompok genom. Konsentrasi ini dibandingkan dengan konsentrasi dari buah ekuivalen yang dimatangkan secara artifisial. Terlepas dari kematangannya, tidak ada perbedaan yang diamati pada daging buah, kulit, atau biji (bila tersedia) dari kultivar mana pun (Gambar S2 dan S3 ). Hal ini menunjukkan bahwa kandungan mineral pada kulit pisang, daging buah, dan biji tetap stabil pascapanen dan sejalan dengan temuan oleh Aquino et al. ( 2014 ), yang melaporkan tidak ada perbedaan signifikan dalam kandungan zat besi antara pisang mentah dan matang di 15 kultivar. Akibatnya, analisis lebih lanjut dilakukan hanya pada buah FG.
Bahasa Indonesia: Untuk memahami hubungan antara pengangkutan mineral dari sumber ke tempat penyerapan pada buah pisang, konsentrasi zat besi dan seng dianalisis dan dibandingkan dalam tiga jenis jaringan buah, yaitu daging buah, kulit, dan biji (bila tersedia). Data yang dirata-ratakan di semua kultivar mengungkapkan tren umum: Kedua mineral ditemukan dalam konsentrasi lebih tinggi pada biji, diikuti oleh kulit, dan terendah pada daging buah (Gambar 2A,B ). Secara khusus, daging buah mengandung konsentrasi zat besi rata-rata 10,2 mg/kg DW, berbeda dengan 29,4 mg/kg pada kulit dan 38,0 mg/kg pada biji (Gambar 2A ). Tren serupa diamati untuk seng, dengan konsentrasi yang jauh lebih tinggi pada kulit dan biji (Gambar 2B ). Karena peran masing-masing sebagai jaringan yang aktif secara fotosintesis atau sebagai organ penyimpanan nutrisi, kulit dan biji diperkirakan mengandung konsentrasi zat besi dan seng yang lebih tinggi, yang dikonfirmasi oleh analisis kami. Dari 33 kultivar yang dianalisis, hanya Musa balbisiana var. liar. Lal Velchi mengandung lebih banyak seng dalam daging buahnya daripada dalam kulitnya. Yang terpenting, biji dari kultivar ini mengandung konsentrasi zat besi dan seng tertinggi yang dilaporkan dalam jaringan buah pisang mana pun, yang menunjukkan pentingnya mereka dalam penyerapan zat besi dalam buah. Namun, dengan merata-ratakan data dari kultivar yang tidak terkait di beberapa kelompok genom, menghasilkan variasi signifikan dalam kandungan mineral, yang dengan jelas menunjukkan kurangnya keseragaman dalam komposisi mineral dalam genus Musa .
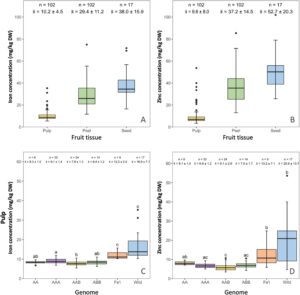
Ketika data bubur buah dianalisis, keempat kultivar liar mengandung konsentrasi zat besi dan seng yang lebih tinggi daripada kelompok genom lainnya dengan rata-rata 16,9 dan 20,8 mg/kg DW, masing-masing (Gambar 2C,D ). Perbedaan dalam konsentrasi zat besi buah ini signifikan ketika dibandingkan dengan semua kultivar yang dapat dimakan kecuali yang dari kelompok Fe’i. Konsentrasi rata-rata seng dalam buah diploid fertil liar berbeda secara signifikan dengan semua kelompok genom kecuali yang dari diploid fertil AA dan kultivar Fe’i. Konsentrasi seng tertinggi diukur dalam bubur Borneo (31,6 mg/kg DW) diikuti oleh Zebrina (27,0 mg/kg DW) dan Lal Velchi (23,0 mg/kg DW) (Tabel 2 ). Malaccensis 845 memiliki konsentrasi seng terendah dari semua kultivar fertil liar yang hanya mengandung 9,1 mg/kg DW, nilai yang mendekati sebagian besar kultivar tanpa biji.
Hanya buah dari Zebrina yang mengandung konsentrasi zat besi di atas “target” pada 28,9 mg/kg DW, diikuti oleh Borneo (16,1 mg/kg DW), Lal Velchi (15,5 mg/kg DW), dan 845 (12,5 mg/kg DW) (Tabel 2 ). Untuk Zebrina, konsentrasi zat besi pulp berkisar antara 20,2 hingga 35,3 mg/kg DW. Variasi ini sebagian dapat dijelaskan oleh kurangnya homogenitas dalam sampel pulp yang dikumpulkan dari kultivar liar. Kehadiran biji dalam kultivar ini menyebabkan pengurangan ukuran mesokarp dan endokarp yang lebih menonjol daripada yang umumnya diamati dari kultivar partenokarpik (Gambar 3A ). Dengan demikian, heterogenitas matriks ini dan kemungkinan kontaminasi kulit biji selama pengambilan sampel mungkin telah menyebabkan variabilitas yang lebih tinggi dalam kandungan zat besi pulp yang terlihat pada kultivar liar. Namun, mengingat peran penting zat besi selama fase heterotrofik awal perkembangan bibit yang berkecambah, konsentrasi tinggi zat besi pulpa dan seng yang diamati pada diploid fertil liar dibandingkan dengan kultivar diploid partenokarpik dan triploid bukanlah hal yang tidak terduga. Zebrina memiliki kandungan zat besi biji tertinggi (61,3 mg/kg DW) dan seng (65,8 mg/kg DW) (Tabel 2 ) sementara Lal Velchi dan 845 memiliki zat besi dan seng terendah, masing-masing. Atas alasan ini, buah dari Zebrina dipilih untuk spektroskopi fluoresensi sinar-X (μ-XRF) dan peta unsur yang digunakan untuk memvisualisasikan lokalisasi zat besi, seng, dan mangan dalam buah Zebrina (Gambar 3 ). Zat besi ditemukan terlokalisasi di dekat dan di sekitar ujung embrio dari bakal biji yang matang sepenuhnya (biji) (Gambar 3B,E ). Secara khusus, sinyal besi tertinggi dideteksi di hilum, mikropil, dan sumbat mikropil pada kulit biji dan area di sekitarnya, sementara sangat sedikit sinyal yang terlihat di endosperma (McGahan 1961 ). Seng didistribusikan secara merata di seluruh ovarium, terutama jaringan pembuluh, epikarp, mesokarp, dan endokarp buah, serta endosperma dan massa kalazal biji (Gambar 3C ). Mangan, yang termasuk di sini sebagai konsentrasi dalam jaringan buah Zebrina melebihi 1000 mg/kg secara kumulatif, terlokalisasi terutama pada dinding epikarp dan karpel, memungkinkan pengamatan yang jelas pada masing-masing lokula dan keseluruhan putik ovarium pisang (Gambar 3D ). Gambar komposit dari ketiga elemen tersebut menunjukkan regulasi besi yang ketat, yang sebagian besar terkonsentrasi di dalam dan di sekitar biji, dan juga mangan yang terlokalisasi pada jaringan septum, sementara seng tidak dikontrol secara ketat, didistribusikan secara difus dalam jaringan buah tanaman pisang (Gambar 3E ). Temuan ini, yang menunjukkan distribusi zat besi yang terlokalisasi dan distribusi seng yang tersebar, konsisten dengan pola yang diamati pada biji padi (Johnson et al. 2011).), dan menyoroti kontrol yang cermat terhadap homeostasis zat besi pada pisang, mungkin sebagai tindakan perlindungan terhadap efek berbahaya spesies oksigen reaktif (ROS) pada kesehatan tanaman (Arosio et al. 2009 ). Meskipun demikian, pisang partenokarpik seperti Cavendish tidak memiliki kapasitas penyerapan seperti biji, yang mungkin menjelaskan konsentrasi zat besi yang lebih rendah yang ditemukan pada kultivar ini.

Kelompok pisang Pacific Fe’i merupakan akumulator zat besi dan seng buah tertinggi berikutnya, dengan konsentrasi rata-rata masing-masing 12,2 dan 13,2 mg/kg DW (Gambar 2B,C ). Konsentrasi zat besi pulpa berbeda secara signifikan ( p < 0,01) dengan semua kelompok genom yang dapat dimakan lainnya (Gambar 2A ). Karena variasi konsentrasi seng pulpa yang lebih tinggi pada kultivar-kultivar ini, perbedaan signifikan hanya diamati pada kultivar AAA ( p < 0,01), AAB ( p < 0,001) dan ABB ( p < 0,05) (Gambar 2B ). Meskipun hanya satu replikasi biologis yang tersedia untuk kultivar Taweyawa dan Karat, yang pertama memiliki konsentrasi zat besi pulpa buah tertinggi ketiga (15,7 mg/kg DW) dan konsentrasi seng tertinggi keempat (24,3 mg/kg DW). Buah Karat mengandung konsentrasi yang sama, dengan zat besi (14,4 mg/kg DW) menempati peringkat kelima dan seng (24,8 mg/kg DW) menempati peringkat ketiga jika dibandingkan dengan semua kultivar yang dianalisis (Tabel 2 ). Satu-satunya kultivar pisang yang dapat dimakan yang menjanjikan yang memenuhi 50% zat besi EAR untuk wanita yang tidak hamil adalah Karat dan Taweyawa. Buah dari kultivar ini juga memiliki konsentrasi seng yang jauh melebihi nilai apa pun yang sebelumnya dilaporkan pada pisang yang dapat dimakan (Davey et al. 2009 ). Kedua kultivar ini serta Asupina adalah satu-satunya pisang partenokarpik yang memiliki daging buah dengan konsentrasi zat besi dan seng yang lebih tinggi daripada kebanyakan EAHB. Namun, harus dicatat bahwa buah dari kultivar ini hanya dianalisis satu kali. Terlepas dari itu, kultivar Fe’i secara konsisten mengungguli rekan-rekan diploid dan triploid mereka (AA, AAA, AAB, ABB) dalam hal kandungan nutrisi, menghasilkan buah dengan konsentrasi zat besi dan seng yang lebih tinggi. Meskipun sangat spekulatif, profil nutrisi superior ini mungkin berasal dari genotipe “T” mereka yang berbeda ( Musa troglodytarum L). Meskipun data tentang kandungan mineral buah kultivar Fe’i terbatas, jelas mereka memiliki jumlah karotenoid yang jauh lebih tinggi daripada kultivar AAA, seperti Cavendish (Buah et al. 2016 ). Akumulasi nutrisi yang meningkat ini diyakini sebagian disebabkan oleh peningkatan kekuatan penyimpanan dalam kultivar ini, yang difasilitasi oleh lebih banyak plastida dalam daging buah (Buah et al. 2016 ). Akibatnya, ini mungkin juga menjelaskan peningkatan konsentrasi zat besi dan seng dalam daging buah ini, sedangkan kandungan mineral kulitnya tetap sebanding dengan kultivar lain yang dianalisis di berbagai genotipe. Konsentrasi zat besi dan seng pada daging buah dalam kelompok genom pisang AAA dari Australia, yang berisi kultivar Cavendish yang populer, rata-rata sebesar 9,1 dan 6,8 mg/kg DW (Gambar 2C,D ). Set ini juga berisi sampel Cavendish Kerdil dan Bogoya (umumnya dikenal sebagai Gros Michel) dari Uganda dengan data yang serupa (Tabel 2 ). Paka dan Terema adalah satu-satunya dua perwakilan dari kelompok diploid yang dapat dimakan (AA) dengan buah yang mengandung rata-rata 8,3 dan 8,1 mg/kg DW zat besi dan seng, masing-masing (Gambar 2C,D ). Dalam kelompok AAB dan ABB, kultivar dengan konsentrasi zat besi tertinggi, Gonja Nakatansese dan Kivuvu, masing-masing, berasal dari Uganda (Tabel 2 ), sementara set Australia rata-rata sebesar 7,8 dan 8,4 mg/kg, masing-masing (Tabel 2 dan Gambar 2C ). Terakhir, kelompok EAHB yang sampel bubur buahnya dalam jumlah besar telah dikumpulkan dari berbagai pertanian di Uganda, rata-rata memiliki 10,7 dan 7,7 mg/kg zat besi dan seng (Tabel 2 ). Analisis konsentrasi zat besi pada kulit menunjukkan bahwa, ketika dirata-ratakan sebagai kelompok, ABB diikuti oleh kultivar liar umumnya mengakumulasikan konsentrasi yang lebih tinggi dalam jaringan ini daripada plasma nutfah lainnya, dengan kultivar AAA mengakumulasikan paling sedikit. Namun, uji Kruskal–Wallis H menunjukkan bahwa tidak satu pun dari perbedaan ini yang signifikan ( p > 0,05) kecuali untuk perbandingan antara kulit kultivar ABB dan AAA (Gambar S4A ). Demikian pula, tidak ada perbedaan signifikan yang diamati dalam konsentrasi seng pada kulit di seluruh kelompok. Konsentrasi seng rata-rata pada kulit berkisar antara 35,0 hingga 40,9 mg/kg DW dan tidak bervariasi secara signifikan antara kelompok genom ( p > 0,05) (Gambar S4B ).
Ada beberapa penelitian pada gandum yang dibiofortifikasi dan dibiakkan secara konvensional (Fan et al. 2008 ; Garvin et al. 2006 ) dan padi (Goloran et al. 2019 ; Moreno-Moyano et al. 2016 ; Impa et al. 2013 ) yang melaporkan penurunan hasil panen seiring dengan peningkatan produksi bahan kering. Fenomena ini sering disebut sebagai “efek pengenceran,” dan kemungkinan didorong oleh dinamika sumber/penyerap yang krusial dalam tanaman (Moreno-Moyano et al. 2016 ). Korelasi dan korelogram Pearson digunakan untuk lebih memahami bagaimana zat besi dan seng dalam buah pisang berpotensi memengaruhi ukuran buah dan pada akhirnya hasil panen (Gambar 4 ). Dalam perbandingan di seluruh kultivar yang dianalisis, konsentrasi zat besi dalam pulp berkorelasi positif signifikan dengan seng ( r = 0,75, p < 0,01) (Gambar 4A ). Korelasi positif ini diamati dalam semua genotipe yang diuji tetapi hanya signifikan untuk kultivar Fe’i ( r = 0,87, p < 0,001) (Gambar 4F ). Lebar dan panjang jari digunakan bersama-sama sebagai indikator berat tandan dengan kedua atribut ini berkorelasi positif signifikan satu sama lain di semua kultivar ( r = 0,71, p < 0,01) dan dengan berbagai tingkat kekuatan dan signifikansi untuk setiap kelompok genom (Gambar 4 ). Secara umum, kedua metrik berkorelasi negatif dengan kandungan zat besi dan seng di seluruh buah (daging buah dan kulit) meskipun hanya zat besi pada daging buah yang tampaknya memiliki pengaruh signifikan (Gambar 4A ). Pengamatan ini tidak tercermin pada tingkat genotipe di mana kandungan zat besi dan seng pada kulit buah tampaknya memainkan pengaruh negatif yang jauh lebih kuat pada ukuran buah terutama pada kultivar AA, Fe’i dan liar (Gambar 4B–G ). “Efek pengenceran” dapat menjelaskan konsentrasi zat besi yang berkurang yang diukur di semua kultivar pisang yang dapat dimakan. Seperti banyak tanaman pangan lainnya, kemungkinan besar tekanan selektif yang disebabkan manusia selama berabad-abad untuk mendapatkan hasil panen yang lebih tinggi pada pisang telah mengorbankan kepadatan nutrisi.

3.3 Profil Mineral yang Berbeda pada Pisang Liar dan Pisang Fe’i Menunjukkan Potensi Biofortifikasi Nutrisi yang Lebih Baik
Untuk menyelidiki apakah terdapat kandungan nutrisi yang lebih tinggi pada kultivar pisang Fe’i dan pisang liar, sembilan mineral esensial yang diukur dalam daging buah semua kultivar tersebut menjadi sasaran analisis komponen utama (PCA). Biplot PCA mencakup 55,4% dari total variasi dalam populasi (Gambar 5A ) dengan pengelompokan dan pemisahan kultivar menjadi tiga kelompok utama, yaitu diploid liar, Fe’i, dan kultivar yang dapat dimakan lainnya (AA, AAA, AAB, dan ABB).

Konsentrasi mineral berkorelasi positif dengan kultivar liar atau Fe’i yang menunjukkan profil konsentrasi mineral unik (ionom) dalam kelompok ini. Secara khusus, zat besi, seng, kalsium, magnesium, natrium, dan mangan sangat terkait dengan kultivar liar dan berkorelasi negatif dengan panjang dan lebar jari. Sebaliknya, tembaga, sulfur, fosfor, dan kalium serta ukuran buah menunjukkan hubungan yang lebih kuat dengan kelompok Fe’i (Gambar 5A ). Kultivar AA, AAA, AAB, dan ABB yang dapat dimakan berkorelasi negatif dengan semua konsentrasi mineral pulp tetapi berkorelasi positif dengan ukuran buah (Gambar 5A ). Analisis ini menunjukkan bahwa diploid liar dan kultivar Fe’i dibedakan oleh ionom yang berbeda dengan pengaruh yang kuat pada ukuran buah. Hal ini dapat dikaitkan dengan peningkatan kekuatan penyimpanan yang dilaporkan pada pisang Fe’i (Buah et al. 2016 ) dibandingkan dengan rekan-rekan genom triploid A dan/atau B partenokarpik mereka, mungkin karena genom T mereka yang unik. Hasil ini mendukung kemungkinan peningkatan profil nutrisi pisang melalui biofortifikasi dengan menggabungkan gen homeostasis besi dari kultivar Fe’i atau diploid fertil. Meskipun demikian, interaksi rumit homeostasis logam pada tanaman menunjukkan bahwa respons multifaktorial yang kompleks mungkin menyertai peningkatan konsentrasi mineral dari ekspresi gen baru (Briat et al. 2015 ; Antosiewicz et al. 2014 ). Yang paling penting adalah kemungkinan kendala hasil yang dapat muncul pada galur transgenik dengan konsentrasi besi yang ditingkatkan.
3.4 Perbandingan Kultivar Musa Australia dan EAHB
Statistik estimasi digunakan untuk menentukan kultivar mana yang tumbuh di Australia memiliki konsentrasi zat besi buah yang secara signifikan lebih tinggi daripada EAHB yang dibudidayakan di Uganda. Plot Gardner–Altman bootstrap digunakan untuk menghasilkan statistik estimasi, dan batas atas interval kepercayaan yang dikoreksi bias dan dipercepat (BCa) untuk konsentrasi zat besi pulp pada kultivar EAHB yang tumbuh di Uganda dihitung menjadi 12,1 mg/kg DW (Gambar 5B ). Batas atas BCa ini menunjukkan bahwa rata-rata populasi sebenarnya untuk konsentrasi zat besi pulp EAHB tidak mungkin melebihi ambang batas ini. Oleh karena itu, kultivar dengan kandungan zat besi di atas 12,1 mg/kg DW dianggap memiliki konsentrasi zat besi pulp yang secara signifikan lebih tinggi daripada EAHB yang tumbuh di Uganda. Dari analisis ini, disimpulkan bahwa, rata-rata, semua kultivar berbiji liar dan tiga kultivar Fe’i menunjukkan konsentrasi zat besi pulp yang secara signifikan lebih tinggi dibandingkan dengan sebagian besar EAHB.
3.5 Identifikasi dan Keanekaragaman Gen Homeostasis Besi pada Musa
Karakterisasi molekuler gen kunci yang terlibat dalam penyerapan, translokasi, remobilisasi, sekuestrasi, dan penyimpanan zat besi sebagian besar diabaikan dalam Musa . Sebagai langkah awal untuk mengatasi masalah ini dan mengidentifikasi target potensial untuk eksploitasi di masa mendatang, identifikasi sistematis gen yang terkait dengan proses ini dilakukan dengan fokus awal pada genom referensi haploid ganda (AA) dari M. acuminata ssp. malaccensis var. Pahang (DHP). Analisis kemudian diperluas ke empat spesies Musa yaitu, Cavendish cv. Baxijiao (Cav, sebagai perwakilan triploid dari kelompok genom acuminata ‘A’), M. balbisiana var. Pisang Klutuk Wulung (PKW, sebagai perwakilan diploid dari kelompok genom balbisiana “B”), dan M. troglodytarum cv. Karat dan Utafan (Kar dan Uta, sebagai perwakilan triploid dan diploid dari kelompok genom troglodytarum “T,” masing-masing). Urutan ortolog diidentifikasi secara serupa dari dua spesies kerabat dekat Musa , Ensete ventricosum (Ev) dan E. glaucum (Eg) untuk digunakan sebagai kelompok luar dalam analisis filogenetik berikutnya (Tabel S5 ). Dalam DHP, urutan 37 gen homeostasis besi diidentifikasi dan posisi kromosomnya dicatat (Gambar 6A dan Tabel S4 ). Di tujuh kultivar, jumlah urutan ortolog yang serupa diidentifikasi, dengan beberapa pengecualian (Tabel S4 ), menghasilkan total 186 urutan gen unik dari Musa dan 75 dari Ensete . Dalam Musa , 14 urutan mengandung beberapa kodon stop atau memiliki kurang dari 70% identitas berpasangan dengan ortolog DHP mereka, dengan 12 di antaranya ditemukan dalam rakitan Fe’i (Tabel S4 ). Akibatnya, perakitan ini menunjukkan frekuensi tertinggi protein “nonfungsional”, yang mungkin mencerminkan keterbatasan dalam penggunaan pembacaan panjang PacBio yang rawan kesalahan dan menyoroti perlunya upaya resequencing (Mitsuhashi et al. 2019 ; Weirather et al. 2017 ; Rhoads dan Au 2015 ). Sebaliknya, peristiwa duplikasi genom utuh (WGD) yang terjadi pada pisang (D’Hont et al. 2012 ) cenderung tidak stabil dan berkembang menuju keadaan diploid melalui penataan ulang struktural dan penyusutan genom, termasuk hilangnya gen duplikat setelah peristiwa ini (Wendel 2015) .). Fragmentasi yang melibatkan penghapusan gen yang terduplikasi dan redundan secara fungsional melalui akumulasi mutasi dapat memainkan peran utama dalam evolusi kelompok Fe’i. WGD merupakan fenomena evolusi yang sangat dominan pada tumbuhan (Van de Peer et al. 2017 ) dengan duplikasi gen dipahami sebagai sumber yang kaya akan hal-hal baru fenotipik (Flagel dan Wendel 2009 ).

Padahal sebagian besar spesies menunjukkan jumlah ortolog yang sebanding dengan DHP, namun Fe’i Uta diploid memiliki dua penghapusan gen ( VIT1a dan VIT4 ), yang berpotensi menunjukkan bahwa paralog ini telah menjadi redundan secara fungsional dan secara progresif telah dihapus dari genom ini. Di sisi lain, duplikasi FRO2a dan FRO5 di Uta dan VIT3 di Kar dapat memberikan beberapa keuntungan evolusi. Peristiwa ini konsisten dengan bentuk neofungsionalisasi (yaitu, divergensi fungsional salinan gen) atau subfungsionalisasi (pembagian ulang ruang-temporal fungsi gen) yang dapat memberikan keuntungan evolusi (Cenci et al. 2019 ). Analisis kami menunjukkan bahwa banyak keluarga gen yang dipelajari memiliki jumlah anggota gen yang lebih tinggi daripada yang dilaporkan sebelumnya dari spesies tanaman lain atau setidaknya menunjukkan bukti duplikasi gen. Hal ini sejalan dengan laporan tentang keberadaan gen duplikat yang substansial dalam genom Musa , khususnya dalam keluarga gen yang diteliti dengan baik, dibandingkan dengan spesies lain (Cenci dan Rouard 2017 ; Cenci et al. 2014 ; D’Hont et al. 2012 ). Lebih jauh, hal ini konsisten dengan tiga WGD yang dilaporkan yang terjadi secara independen dari garis keturunan monokotil lainnya selama sejarah evolusi pisang, menghasilkan genom paleopoliploid kompleks dengan beberapa paralog (D’Hont et al. 2012 ).
Sementara perluasan sebagian besar keluarga gen homeostasis besi diamati, ada beberapa pengecualian. Misalnya, hanya empat ortolog gen NAS yang ditemukan pada pisang, yang sama seperti pada Arabidopsis (Klatte et al. 2009 ; Bauer et al. 2004 ) dan hanya satu lebih banyak daripada yang dilaporkan dari padi (Johnson et al. 2011 ). Hanya dua gen IRT yang diidentifikasi di seluruh genom Musa yang lebih luas dalam studi ini, yang sangat mirip dengan studi yang dilaporkan dari padi (Ishimaru et al. 2007 ; Bughio et al. 2002 ) dan Arabidopsis (Vert et al. 2002 ; Eide et al. 1996 ) di mana mereka telah dikarakterisasi dengan baik. Delapan gen FRO telah didokumentasikan dalam Arabidopsis (Mukherjee et al. 2006 ) sementara hanya dua yang telah diidentifikasi dalam padi. Namun, tidak satu pun dari ini menunjukkan aktivitas reduktase Fe 3+ -chelate yang signifikan ketika diuji dalam ragi, mungkin mencerminkan karakteristik tanaman Strategi II (Li et al. 2019 ). Pemeriksaan ekstensif genom Arachis hypogaea (kacang tanah) mengungkapkan bahwa AhFRO mengalami duplikasi WGD/segmental, yang mengakibatkan perluasan famili gen AhFRO menjadi sembilan gen, lima di antaranya adalah duplikasi AhFRO2 (Guan et al. 2024 ). Dalam semua genom pisang yang diselidiki, FRO2 ditemukan terduplikasi setidaknya sekali pada kromosom yang sama, sebuah pengamatan yang juga terlihat pada kedua spesies Ensete yang dianggap sebagai kerabat jauh Musa . Kecuali Uta, yang mengalami peristiwa duplikasi tambahan FRO2a dan FRO5 , lima gen FRO diidentifikasi di antara spesies Musa yang dianalisis . Duplikasi ini dapat dikaitkan dengan sifat allopoliploid spesies Musa , meskipun mungkin ada keuntungan evolusi dalam menghindari hilangnya gen, terutama karena pisang merupakan strategi penanaman (Römheld 1987b ) yang bergantung pada FRO2 untuk penyerapan zat besi yang efisien dari tanah. Peristiwa duplikasi yang diamati dalam famili gen feritin pisang, khususnya Fer3 , mengarah pada identifikasi lima gen feritin, melampaui yang ditemukan di Arabidopsis sebanyak satu (Petit et al. 2001 ) dan padi sebanyak tiga (Nguyen et al. 2022 ; Gross et al. 2003 ). Demikian pula, spesies Musa menunjukkan peristiwa duplikasi dalam VITgen keluarga, terutama pada Kromosom 11, dan duplikasi VIT1 pada Kromosom 7 dan 9. Kultivar Kar juga merupakan satu-satunya spesies Musa yang memiliki gen VIT3 tambahan yang ditemukan pada kromosom yang sama. Sebuah studi tentang evolusi molekuler gen keluarga VIT mengungkapkan berbagai macam nomor gen VIT pada tanaman, mulai dari 2 hingga 21, dengan contoh pertambahan dan kehilangan (Cao 2019 ). Selain itu, gandum heksaploid mengandung 31 gen pengangkut besi seperti vakuolar ( VTL ) (Sharma et al. 2020 ), sementara Arabidopsis memiliki satu gen VIT (Kim et al. 2006 ) dan dua gen VTL yang diketahui (Gollhofer et al. 2014 ). Sebaliknya, padi hanya mengandung dua gen VIT (Zhang et al. 2012 ). Mengingat sifat rumit dari keluarga gen VIT dan homologi sekuens signifikan yang dimiliki oleh gen VIT dan VTL (Gollhofer et al. 2014 ), masuk akal bahwa beberapa gen VIT yang teridentifikasi di Musa sebenarnya adalah gen VTL atau sekadar duplikasi gen VIT lainnya . Oleh karena itu, penyelidikan lebih lanjut terhadap keluarga gen VIT Musa diperlukan.
Keluarga gen YSL menunjukkan keragaman yang signifikan di seluruh spesies tanaman sambil mempertahankan struktur protein transmembran yang sangat terkonservasi (Chowdhury et al. 2022 ). Arabidopsis hanya memiliki delapan gen pengkode protein YSL (Hofstetter et al. 2013 ), sedangkan 13 ditemukan pada spesies Musa yang diperiksa dalam penelitian ini. Tanaman monokotil, secara umum, telah mengalami peristiwa duplikasi umum dalam keluarga gen YSL , yang mengakibatkan perluasan (Chowdhury et al. 2022 ). Misalnya, padi (Aoyama et al. 2009 ; Koike et al. 2004 ) dan jagung (Song et al. 2024 ) masing-masing mengekspresikan 18 dan 19 transporter YSL yang berbeda. Duplikasi ini diperkirakan memainkan peran penting dalam diversifikasi fungsional keluarga gen YSL dengan mengubah pola ekspresi gen spasial dan temporal dan sebagian menjelaskan tingginya jumlah gen YSL yang ditemukan dalam pisang (Chowdhury et al. 2022 ).
Secara keseluruhan, duplikasi gen kunci dalam famili protein Fer, FRO, VIT, dan YSL diamati pada semua spesies Musa yang diteliti, dengan kultivar Fe’i menunjukkan jumlah kejadian duplikasi tertinggi. Temuan ini menyiratkan upaya terpadu dalam evolusi Musa untuk mempertahankan gen yang penting bagi perolehan, pengangkutan, dan penyerapan zat besi. Menariknya, kultivar pisang yang mengandung genom T tidak dapat disangkal menunjukkan peningkatan efisiensi dalam proses biologis ini dibandingkan dengan kultivar pisang yang mengandung genom A dan B yang lebih umum dan lazim secara komersial sebagaimana dibuktikan oleh kandungan zat besi yang tinggi yang diamati dalam buah spesies Fe’i yang berbeda.
3.6 Analisis Filogenetik Gen Homeostasis Besi pada Musa
Analisis filogenetik yang menggabungkan sekuens dari kerabat liar dapat menawarkan wawasan berharga ke dalam hubungan evolusi di antara taksa yang dipelajari, menjelaskan divergensi genetik, sejarah filogenetik, dan perolehan sifat atau adaptasi unik tertentu (Brozynska et al. 2016 ). Oleh karena itu, analisis filogenetik dilakukan dengan tujuan untuk memahami bagaimana protein pengatur zat besi mungkin berevolusi dari diploid berbiji leluhur (AA dan BB) dan kultivar Fe’i, dan bagaimana genom ini berhubungan dengan kultivar triploid partenokarpik modern seperti Cavendish. Sekuens gen yang teridentifikasi diterjemahkan menjadi protein, dan untuk kejadian duplikat, sekuens dengan identitas berpasangan tertinggi dengan referensi DHP dipilih untuk analisis filogenetik (Tabel S4 ). Protein ini dipertahankan untuk kelengkapan filogenetik. Protein homeostasis besi ortolog dari Ev dan Eg digunakan sebagai kelompok luar, menggantikan protein Ev dengan Eg di mana terdapat beberapa kodon stop atau identitas rendah (Tabel S5 ).
Untuk analisis, keluarga protein (Gambar 6B ) serta perbandingan protein individu (Gambar S6-S9 ) dibuat. Pola pengelompokan yang diamati adalah sama di seluruh keluarga protein, dengan sekuens yang berasal dari dua genom T yang mengandung kultivar Kar dan Uta secara konsisten membentuk klade terpisah yang lebih erat terkait dengan kelompok luar Ensete daripada kultivar lain dan secara konsisten menunjukkan divergensi evolusi yang lebih besar dari kultivar yang mengandung genom A (Cav dan DHP) dibandingkan dengan kultivar PKW yang mengandung genom B (Gambar 6B ). Dalam semua kecuali dua contoh, node ini didukung oleh nilai bootstrap yang melebihi 92%, dengan salah satu pengecualian ini terletak di dalam pohon keluarga IRT, yang hanya berisi dua anggota keluarga dengan sekuens Fe’i yang ambigu. Pola pengelompokan ini juga diamati pada 59,5% (22/37) dari semua protein individu yang diselidiki (Gambar S5-S9 ) dan turun menjadi 37,5% untuk keluarga VIT. Dalam kasus langka di mana protein dari kultivar genom T dan A mengelompok lebih dekat satu sama lain, seperti pada FRO2b (Gambar S6 ), NAS2 (Gambar S7B ), VIT2, dan VIT4 (Gambar S8 ), serta YSL6a, YSL8, dan YSL12e (Gambar S9 ), analisis tersebut mengandung protein nonfungsional dan/atau menunjukkan nilai bootstrapping yang rendah.
Analisis ini menyoroti fitur-fitur yang terkonservasi dan divergen yang mencerminkan mekanisme adaptasi khusus untuk metabolisme zat besi dalam genus Musa . Secara khusus, analisis ini kembali menunjukkan proteom yang berbeda dan divergen untuk penyerapan dan pengaturan zat besi pada pisang Fe’i (T) dan Cavendish (A). Mengingat hal ini, gen-gen kunci yang terlibat dalam homeostasis zat besi pada pisang Fe’i harus divalidasi secara fungsional dengan tujuan untuk mengeksploitasinya sebagai kandidat cisgen untuk biofortifikasi zat besi. Untuk memfasilitasi hal ini, analisis ekspresi diferensial dari semua gen yang diidentifikasi sebelumnya dilakukan pada jaringan akar, daun, dan buah dari kultivar pisang Cavendish “Grande Naine” dan “Williams.”
3.7 Ekspresi Diferensial Gen Homeostasis Besi pada Cavendish
Ekspresi gen diferensial dan variasi fenotipik yang dihasilkan dalam spesies Musa sering kali dapat dikaitkan dengan subgenomnya yang berbeda. Misalnya, kultivar genom B telah menunjukkan peningkatan regulasi pada gen yang terkait dengan biosintesis flavonoid, metabolisme asam lemak, serta metabolisme gula amino dan nukleotida (Cenci et al. 2019 ). Selain itu, penataan ulang kromosom ganda dan variasi sekuens dalam genom T triploid Kar dan genom T diploid Uta telah menghasilkan sifat buah khas yang terkait dengan peningkatan kualitas nutrisi, termasuk pematangan nonklimakterik dan peningkatan konsentrasi karotenoid, riboflavin, flavonoid, dan galaktosa bebas (Zhou et al. 2024 ; Li et al. 2022 ).
Pendekatan kami dimulai dengan menyelidiki ekspresi gen di Cavendish dalam kondisi yang cukup zat besi untuk mengidentifikasi hambatan dasar. Untuk mencapai hal ini, kumpulan data RNA-seq dari jaringan akar, daun, dan buah Cavendish dipetakan secara sistematis ke 37 gen homeostasis zat besi yang diidentifikasi dari perakitan DHP. Pola ekspresi relatif dibandingkan di seluruh jaringan ini (Gambar 6C ) untuk memfasilitasi pemilihan gen kandidat untuk biofortifikasi dan untuk meningkatkan pemahaman kita tentang pola ekspresi spasial gen-gen ini di dalam tanaman.
Gen yang mengkode feritin ( Fer ) ditemukan diekspresikan terutama di daun dan buah, dengan Fer2 dan Fer3b diekspresikan secara diferensial paling kuat di buah, sementara Fer1 , Fer4 , dan Fer3a memiliki ekspresi diferensial yang lebih tinggi di daun (Gambar 6C ). Di antara ini, Fer3b , gen duplikat, menunjukkan tingkat ekspresi keseluruhan tertinggi di buah, yang lebih dari lima kali lipat lebih tinggi daripada gen Fer tertinggi berikutnya ( Fer1 di daun). Fer3b juga ditemukan diekspresikan secara substansial di daun (13.502 hitungan bacaan yang dinormalisasi) jika dibandingkan dengan Fer1 , yang merupakan gen yang diekspresikan daun tertinggi dengan 37.483 hitungan bacaan yang dinormalisasi. Fer3 diyakini memiliki peran yang mirip dengan gen AtFer2 Arabidopsis , yang diketahui menyimpan zat besi khususnya dalam biji (Petit et al. 2001 ) dan telah muncul sebagai gen kandidat kuat untuk penyimpanan zat besi dalam pisang, yang kemungkinan memainkan peran penting tidak hanya selama pematangan buah tetapi juga pada tahap perkembangan lainnya.
Gen oksidase ferri reduktase kloroplas, FRO7 dan FRO8 , memiliki ekspresi diferensial yang kuat pada jaringan daun dibandingkan dengan jaringan akar dan buah, di mana ekspresi kedua gen tersebut minimal (Gambar 6C ). Pola ini selaras dengan temuan di Arabidopsis, di mana AtFRO7 dan AtFRO8 sebagian besar diekspresikan dalam jaringan fotosintesis dan pucuk, masing-masing (Mukherjee et al. 2006 ). Tingkat ekspresi rendah FRO2a dan FRO5 tetapi juga IRT1 dan IRT2 diamati di semua jaringan pisang, dengan ekspresi tertinggi terdeteksi di jaringan akar (Gambar 6C ). Pola ekspresi ini mencerminkan ortolog Arabidopsis masing-masing, AtIRT1 (Vert et al. 2002 ), AtIRT2 (Vert et al. 2009 ), AtFRO2 , dan AtFRO5 (Mukherjee et al. 2006 ), yang diketahui diekspresikan di akar. Ekspresi rendah gen-gen ini diharapkan terjadi pada kondisi yang cukup zat besi, yang menunjukkan penyerapan zat besi yang cukup dari tanah pada tanaman dewasa. Namun, mengingat pentingnya gen-gen ini untuk penyerapan zat besi pada tanaman monokotil non-graminaceous seperti pisang, peningkatan regulasi yang kuat diharapkan terjadi pada kondisi kekurangan zat besi, seperti halnya pada Arabidopsis (Vert et al. 2002 ) dan padi (Lee dan An 2009 ).
Gen NAS yang teridentifikasi tampaknya diekspresikan secara dominan di akar dan pada tingkat yang jauh lebih tinggi dibandingkan dengan jaringan lain (Gambar 6C ). NAS4 menunjukkan ekspresi keseluruhan tertinggi, yang tiga kali lipat lebih tinggi daripada gen tertinggi berikutnya, NAS2 . NAS3 ditemukan sebagai satu-satunya gen NAS yang diekspresikan dalam buah pisang, meskipun perkiraan jumlah bacaannya konsisten dengan aktivitas rendah dalam jaringan ini (Gambar 6C ). Karena ekspresi NAS terutama terbatas pada akar, sintesis nikotinamina (NA) kemungkinan terjadi pada akar pisang, dan kemudian NA, yang mungkin terikat pada zat besi, diangkut ke bagian lain tanaman sebagaimana diperlukan. Pola lokalisasi ini selaras dengan temuan pada padi, di mana NAS sebagian besar terlokalisasi pada vesikel dalam sel akar yang kekurangan zat besi, memungkinkan sintesis NA yang efisien dan memfasilitasi penyerapan zat besi dan transportasi jarak jauh di dalam tanaman (Kobayashi dan Nishizawa 2012 ). Demikian pula pada jagung, ZmNAS1 dan ZmNAS2 diekspresikan dalam korteks dan stele akar, sementara ZmNAS3 biasanya diekspresikan dalam meristem aksiler, primordia daun, dan sel mesofil (Zhou et al. 2013 ). Pada pisang, ekspresi NAS1 tidak terdeteksi, sementara NAS3 diekspresikan secara eksklusif dalam buah tetapi dalam jumlah yang rendah. Mengingat jumlah ekspresi yang sangat rendah yang terdeteksi dari gen-gen ini dalam jaringan yang diuji, ada kemungkinan bahwa organ atau jaringan yang lebih terspesialisasi berfungsi sebagai tempat utama ekspresi.
Secara umum, delapan gen VIT yang diidentifikasi memiliki ekspresi rendah di sebagian besar jaringan yang diuji, kecuali VIT1b dengan ekspresi buah 40 kali lipat lebih tinggi daripada gen VIT lainnya di semua jaringan yang diuji (Gambar 6C ). VIT4 memiliki tingkat ekspresi yang terukur di daun (Gambar 6C ) sementara itu juga memiliki tingkat ekspresi yang sebanding di jaringan akar yang mirip dengan gen VIT5 dan VIT1b . Ekspresi rendah gen VIT di Musa dapat menunjukkan bahwa (i) pada saat pengambilan sampel tanaman telah menyimpan sebagian besar zat besi yang diserapnya atau (ii) gen yang diduplikasi ini telah menjadi berlebihan. Ekspresi buah yang kuat dari VIT1b menunjukkan peran yang mirip dengan AtVIT1 yang terlokalisasi secara spesifik pada vakuola biji Arabidopsis (Kim et al. 2006 ). Pola ekspresi ini menunjukkan bahwa Cavendish VIT4 dapat berfungsi mirip dengan gen OsVIT1 dan OsVIT2 padi , yang paling aktif di daun bendera (Zhang et al. 2012 ). Penelitian lebih lanjut diperlukan untuk mengeksplorasi potensi paralel fungsional ini pada pisang. Peningkatan regulasi VIT1b yang signifikan pada buah dan kurangnya peningkatan penyimpanan zat besi terkait pada jaringan tersebut masih belum dapat dijelaskan dan juga memerlukan penyelidikan. Dari perspektif biofortifikasi dan dengan keadaan pengetahuan saat ini, VIT1b mungkin bukan kandidat yang ideal untuk meningkatkan penyimpanan zat besi pada buah. Sebaliknya, VIT4 yang menunjukkan ekspresi sedang pada daun pisang, tempat akumulasi zat besi yang diketahui, dapat menjadi kandidat yang lebih cocok untuk meningkatkan konsentrasi zat besi pada buah jika diekspresikan dengan cara yang spesifik pada buah.
Gen YSL , keluarga gen terbesar yang dianalisis dalam studi ini, menunjukkan variasi terluas dalam ekspresi gen di ketiga jenis jaringan pisang. Ini sejalan dengan temuan pada jagung, di mana gen YSL terbukti diekspresikan dalam beberapa jaringan (Song et al. 2024 ). Pada pisang, ekspresi gen YSL keseluruhan tertinggi diamati pada buah, kemungkinan dipengaruhi oleh tahap perkembangannya selama pengambilan sampel. Klade yang terdiri dari enam gen YSL ditemukan diekspresikan secara berbeda dalam buah Cavendish, dengan YSL12c memiliki ekspresi terkuat sejauh ini lebih dari 4,5 kali lipat dari YSL12f , gen yang diekspresikan paling tinggi berikutnya dalam klade itu (Gambar 6C ). Ekspresi yang menurun kemudian ditemukan di YSL6a , YSL3 , YSL6b , dan YSL14 . Meskipun ekspresi YSL14 secara berbeda disukai terhadap buah, perkiraan jumlah bacaan dalam jaringan ini serta dua jenis jaringan lain yang diuji menunjukkan ekspresi yang sangat rendah. Menariknya, dari enam gen dalam klade itu, semua kecuali satu ( YSL6b ) memiliki profil ekspresi jaringan yang sama dengan ekspresi terendah di akar, diikuti oleh daun dan akhirnya buah. Dua gen dalam klade yang berdekatan diekspresikan secara berbeda di akar, dengan YSL12b menjadi ekspresior terkuat dari keduanya. Namun, gen yang paling banyak diekspresikan dalam jaringan akar adalah YSL12c , YSL6a , dan YSL6b . Tiga gen YSL dengan ekspresi daun terkuat adalah YSL12c diikuti oleh YSL12g dan YSL6a (Gambar 6C ). Yang penting, YSL12c , YSL6a , dan YSL3 menunjukkan ekspresi tinggi di ketiga jaringan, yang menunjukkan potensi keterlibatan mereka dalam transportasi jarak jauh kation divalen. Pola ekspresi ini menunjukkan bahwa gen-gen ini dan gen-gen dalam klade yang sama mungkin termasuk dalam Subkelompok I YSL , yang terkait dengan transportasi besi jarak jauh. Dari semua itu, YSL12c menunjukkan ekspresi tertinggi dalam buah, menjadikannya kandidat yang menjanjikan untuk upaya biofortifikasi di masa mendatang. Pada jagung, ZmYSL3 dan ZmYSL11 adalah YSL Subgrup I , sementara beberapa YSL lainnya ( ZmYSL2 , 5 , 7 , 9 , dan 12)) menunjukkan ekspresi yang relatif tinggi dalam embrio atau endosperma (Song et al. 2024 ). Pada tanaman yang diteliti dengan baik seperti Arabidopsis, padi, dan jagung, Subgroup II YSL seperti AtYSL4 dan AtYSL6 (Divol et al. 2013 ), OsYSL6 (Sasaki et al. 2011 ), ZmYSL4 dan ZmYSL5 (Song et al. 2024 ), terutama mengatur penyimpanan dan detoksifikasi logam daripada transportasi jarak jauh. Meskipun identitas spesifik gen-gen Subgroup II YSL ini dalam pisang Cavendish masih belum jelas, ada kemungkinan bahwa gen-gen dalam klade kedua (Gambar 6C ) dapat berpartisipasi dalam detoksifikasi dan penyimpanan logam lokal. Gen-gen ini biasanya menunjukkan ekspresi hanya dalam satu jaringan spesifik, seperti YSL12b , yang sebagian besar diekspresikan dalam akar, atau YSL12g , yang terutama diekspresikan dalam daun. Kemungkinan, kedua klade yang diidentifikasi dalam profil ekspresi gen YSL dapat bersesuaian dengan peran fungsional yang berbeda dari gen YSL Subgrup I dan II pada pisang, sehingga menjelaskan pola ekspresi tumpang tindih yang diamati di berbagai jaringan. Menjelaskan peran gen-gen ini pada pisang sangat penting untuk memahami mekanisme pengaturan yang mengatur keluarga gen penting ini pada tanaman pangan utama.
4 Arah Masa Depan
Studi ini berusaha untuk meletakkan dasar untuk mengidentifikasi dan mengkarakterisasi gen-gen homeostasis besi utama di Musa , yang bertujuan untuk menentukan kandidat cisgene yang cocok untuk tujuan biofortifikasi. Dengan memanfaatkan gen-gen homeostasis besi yang bersumber dari kultivar dengan kandungan besi dan seng yang tinggi secara alami, pendekatan yang disesuaikan untuk meningkatkan profil nutrisi pisang dapat dirancang di masa depan. Hasil penting dari penyaringan plasma nutfah adalah kemampuan akumulasi nutrisi yang unggul dari pisang yang termasuk dalam kelompok Fe’i, dan menggarisbawahi potensi pisang-pisang ini sebagai sumber daya yang berharga dalam upaya biofortifikasi, sebuah konsep yang sebelumnya diilustrasikan oleh Paul et al. ( 2017 ). Pada pisang Fe’i, baik profil ionom pulp dan protein-protein homeostasis besi sangat berbeda dari kultivar-kultivar pisang yang dibudidayakan secara komersial. Akibatnya, melakukan studi ekspresi gen yang komprehensif pada pisang-pisang ini dalam kondisi yang cukup dan kekurangan besi merupakan perkembangan alami untuk upaya-upaya penelitian di masa depan.

